

2 Общая схема строения растительной клетки
Основными, самыми общими компонентами, из которых построены клетки растений, являются ядро, цитоплазма с многочисленными органоидами различного строения и функций, оболочка, вакуоль. Оболочка покрывает клетку снаружи, под ней находится цитоплазма, в ней — ядро и одна или несколько вакуолей. Как строение, так и свойства клеток разных тканей в связи с их разной специализацией резко различаются. Перечисленные основные компоненты и органоиды развиты в них в различной степени, имеют неодинаковое строение, а иногда тот или иной компонент может вовсе отсутствовать. На рисунке 6 представлена схема соотношения между отдельными видами структур во взрослой растительной клетке

Рисунок 6 — Соотношение между отдельными видами структур во взрослой растительной клетке
В отличие от клеток других эукариотических организмов для растительной клетки характерно следующее:
наличие пластидной системы, возникшей в связи с фототрофным способом питания;
жесткая полисахаридная клеточная стенка, окружающая клетку;
центральная вакуоль в зрелых клетках;
отсутствие у делящейся растительной клетки центриолей;
совмещение в себе трех относительно автономных генетических систем: ядерной (хромосомной), митохондриальной и пластидной.
При этом растительная клетка, в отличие от других эукариотических клеток, образовалась как минимум из трех исходно самостоятельных форм:
«хозяйского» организма, генетический аппарат которого переместился в ядро;
гетеротрофной бактерии (похожей на родоспириллу), послужившей предшественницей митохондрий;
древнейшей бактерии с оксигенным фотосинтезом, похожей на цианобактерию Sinechocystis, ставшей родоначальницей пластид.
Длительная коэволюция симбионтов привела к перераспределению функций между ними и их генетическими системами, при этом многие гены митохондриальной и пластидной ДНК были перемещены в ядро.
Рассматривая общие черты строения и жизнедеятельности растительных клеток, удобно говорить о некоей типовой клетке. Такую клетку, хотя она и не существует в «чистом виде», можно изобразить (рисунок 7). Из реальных клеток более других похожи на такую «типовую» клетки паренхимы листа (рисунок 8).
3 Мембраны растительной клетки
Структура. Толщина биомембран не превышает 6-10 нм. Согласно жидкостно-мозаичной гипотезе строения биологических мембран основу мембраны составляет двойной слой фосфолипидов с некоторым количеством других липидов (галактолипидов, стеринов, жирных кислот и др.), причем липиды повернуты друг к другу своими гидрофобными концами (рисунок 9). Участки молекул полярных липидов, образованные очень часто ненасыщенными жирными кислотами, и стерины обеспечивают несколько разрыхленное (жидкое) состояние бислоя. Полярной частью молекула фосфолипида хорошо взаимодействует с водной средой. Кратная связь в довольно длинный неполярном «хвосте» фосфолипидов находится в цис-конфигурации, поэтому «ножка» не прямая, а как бы изогнутая (рисунок 10 а).
Если образец фосфолипида размешать в водной среде, образуются так называемые мицеллы (рисунок 10 б) построенные так, что полярные головы обращены в водный слой, а неполярные хвосты — внутрь мицеллы. В определенных условиях можно получить липосомы, состоящие из бислоя липидов (рисунок 10 г) и очень напоминающие клетку, только в миниатюре. У липосом имеется внутреннее пространство, практически изолированное от внешней среды вследствие того, что структура липосомы чрезвычайно стабильна. Способность некоторых липидов к «самосборке» в двойные слои является очень важным свойством, имеющим решающую роль в построении клеточных мембран.

Рисунок 7 — Обобщённая схема строения растительной клетки, составленная по данным электронно-микроскопического исследования разных растительных клеток:
1— аппарат Гольджи; 2 — свободно расположенные рибосомы; 3 — хлоропласты; 4 — межклеточные пространства; 5 — полирибосомы (несколько связанных между собой рибосом); 6 — митохондрии; 7 — лизосомы; 8 — гранулированная эндоплазматическая сеть; 9 — гладкая эндоплазматическая сеть; 10 — микротрубочки; 11 — пластиды; 12 — плазмодесмы, проходящие сквозь оболочку; 13 — клеточная оболочка; 14 — ядрышко; 15, 18 — ядерная оболочка; 16 — поры в ядерной оболочке; 17 — плазмалемма; 19 — гиалоплазма; 20 — тонопласт; 21 — вакуоли; 22 — ядро.

Рисунок 8 — Клетка из губчатой паренхимы листа коровяка (Verbascum thapsus). Электронная микрофотография (увел. ×25000) М. Ф. Даниловой:
ко — клеточная оболочка; пд — плазмодесмы, соединяющие цитоплазму соседних клеток; мп — межклеточное пространство; ц — цитоплазма; я — ядро; в — вакуоль; яо — ядерная оболочка; м — митохондрии; хл — хлоропласты; пл — плазмалемма; тп — тонопласт.
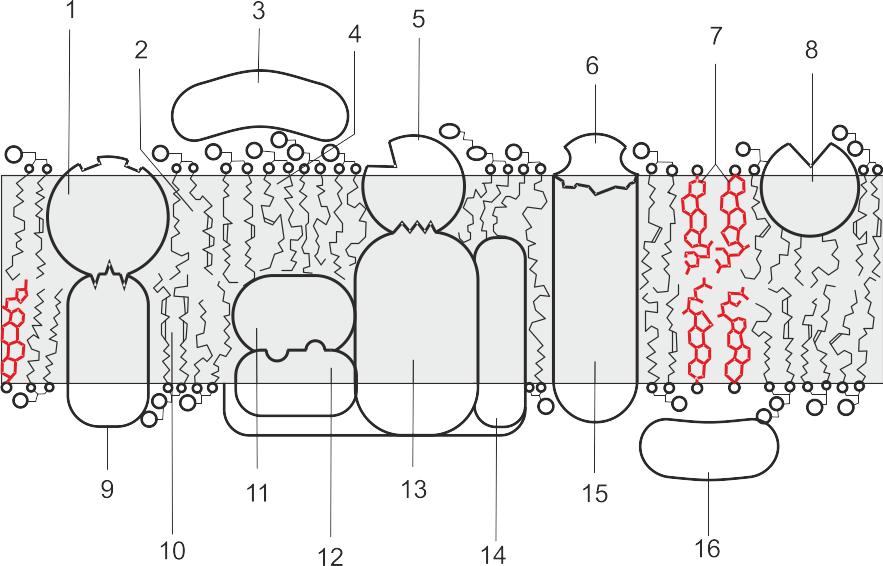
Рисунок 9 — Схема строения цитоплазматической мембраны:
1 — регуляторная субъединица фермента; 2 — фосфатидная кислота; 3 — периферийный белок; 5 — регуляторная субъединица насоса; 6 — регуляторная субъединица канала; 7 — стерины; 8 —переносчик (глобулярный мембранный белок); 9 — фермент; 10 — фосфатид (лецитин и др.); 11 — фермент (например, протеинкиназа); 12 — регуляторная субъединица фермента; 13 — ионный насос; 14 — структурный белок; 15 — ионный канал; 16 — периферийный белок.
|
|
а |
б |
|
|
в |
|



Рисунок 10 — Полярность в расположении молекул фосфолипидов в мембранных структурах: а — молекула фосфолипида; б — мицелла; в — участок плазматической мембраны; г — липосома
Наружная и внутренняя стороны биологических мембран обращены в качественно разные гидрофильные среды. Это является причиной асимметричного строения мембран — в наружном слое плазмалеммы содержится больше стеринов и гликолипидов.
Липиды, входящие в состав мембранного бислоя, не закреплены жестко, а непрерывно меняются местами. Перемещения липидных молекул бывают двух типов:
в пределах своего монослоя (латеральная диффузия);
путем перестановки двух липидных молекул, противостоящих друг другу в двух монослоях («флип-флоп»).
При латеральной диффузии молекулы липидов претерпевают миллионы перестановок в секунду, а скорость ее составляет 5—10 мкм/с. Перестановки молекул липидов из одного монослоя в другой происходят значительно реже, но могут ускоряться мембранными белками.
В состав мембран входят разные в функциональном отношении белки: белки-ферменты, белки, выполняющие функции насосов, переносчиков, ионных каналов, белки-регуляторы и структурные белки (рисунок 9).
Если такие специализированные протеиновые комплексы :
погружены в липидную фазу и удерживаются гидрофобными связями (липопротеины), их называют «интегральные белки»;
удерживаются на внутренней и внешней поверхностях мембран электростатическими связями (гидрофильные белки), взаимодействуя с гидрофильными головками полярных липидов, то их называют «периферийные белки».
Периферийные белки ассоциированы с мембраной за счет присоединения к интегральным белкам или липидному бислою слабыми связями: водородными, электростатическими, солевыми мостиками. Они в основном растворимы в воде и легко отделяются от мембраны без ее разрушения. Некоторые периферийные белки обеспечивают связь между мембранами и цитоскелетом.
Интегральные белки мембран нерастворимы в воде. Как минимум один из доменов интегрального белка встроен в гидрофобную часть бислоя мембраны, поэтому интегральный белок, как правило, не может быть удален из мембраны без ее разрушения. Интегральные белковые глобулы располагаются в фосфолипидных слоях мембран ориентированно. Определенные участки липопротеиновых глобул (участки узнавания) служат для самосборки полиэнзимных мембранных комплексов. На положение белков в мембране оказывают влияние состав фосфолипидов (свободных и связанных), величина электростатического заряда мембраны.
В последнее время показано существование третьей группы белков, так называемых «заякоренных» в мембране белков. Эти белки фиксируются в мембране за счет специальной молекулы, в качестве которой могут выступать жирная кислота, стерин, изопреноид или фосфатидилинозитол. Белки, связанные с изопреноидами (пренилированные белки) или жирной кислотом, обратимо соединяются с эндоплазматической (внутренней) поверхностью мембраны. В отличие от этих двух групп белков фосфатидилинозитолсвязанные белки находятся с внешней (экстрацеллюлярной, или люменальной) стороны мембраны.
Функциональная активность мембран и изменения мембранного потенциала сопровождаются всплыванием или погружением субъединиц, их латеральными перемещениями.
Особенностью липидного состава плазмалеммы по сравнению с другими мембранами растительной клетки является высокое содержание стеринов, но в отличие от плазматической мембраны животной клетки для плазмалеммы характерна высокая вариабельность их состава в зависимости от вида растения, органа и ткани.
Функции мембран
1) Структурная функция. У эукариотической клетки в отличие от прокариотической клетки множество внутренних отсеков (компартментов), окруженных мембранами, и различающихся по степени активности содержащихся в них химических соединений и систем, регулирующих их превращения. Компартменты эукариотической клетки называются органеллами. Таким образом, клеточные мембраны выполняют функцию расчленения биохимических процессов, разделения их между различными компонентами протоплазмы и пространственного размещения в объеме клетки фондов метаболитов и ферментов.
Одномембранные структурные компоненты клеток: плазмалемма, эндоплазматический ретикулум, комплекс Гольджи, лизосомы, вакуоль.
Двухмембранные структурные компоненты клеток: ядро, митохондрии, пластиды.
2) Барьерная функция. Для клеток и субклеточных частиц мембраны служат механическим барьером, отделяющим их от внешнего пространства.
3) Контроль поглощения и секреции веществ (транспортная функция). В плазмалемме находятся множество транспортных систем, прежде всего для транспорта ионов, — ионные каналы, ионные переносчики и ионные насосы. Благодаря им осуществляется очень точный и селективный транспорт ионов как внутрь клетки, так и наружу, т.е. оптимальное снабжение клетки необходимыми ионами.
Через плазмалемму происходит также перенос макромолекул. Так, в периплазматическое пространство транспортируются строительные блоки клеточной стенки — полисахариды и структурные белки. Эти соединения обычно находятся в везикулах аппарата Гольджи и высвобождаются из них путем экзоцитоза.
4) Аккумуляция и трансформация энергии. Плазмалемма любой клетки является энергизованной мембраной, т. е. на ней существует градиент электрохимического потенциала ∆μН+, который используется для выполнения полезной работы, прежде всего для активного переноса веществ через мембрану. В хлоропластах зеленых растений энергия света трансформируется в энергию NADH и АТР, а в конечном счете — в стабильную энергию химических связей сахаров, органических кислот, аминокислот.
5) Размещение и обеспечение работы ферментов. В плазмалемме находится множество ферментов. Это ферменты построения клеточной стенки (целлюлозосинтаза), ферменты сигнальных систем (фосфолипазы С, А; аденилатциклаза) и ряд других ферментов (например, цитохром b5).
6) Рецепторная функция. Плазмалемма клетки — это мозаика различных рецепторов эндогенных сигналов (прежде всего фитогормонов) и внешних воздействий. К последним относятся рецепторы элиситоров (веществ, выделяемых патогенами) и рецепторы физических факторов — температуры, давления и др. Температура изменяет жесткость (текучесть) мембраны, т.е. ее механические свойства. Изменение этих свойств плазмалеммы приводит к открытию или закрытию находящихся в ней механосенсорных ионных каналов. Таким образом, плазмалемма является местом рецепции (восприятия) различных сигналов химической и физической природы.
7) Сигнальная функция. Многие компоненты плазмалеммы после восприятия сигналов служат источником вторичных мессенджеров — молекул, которые «передают» сигнал по эстафете и усиливают его. В качестве вторичных мессенджеров выступают инозитол-1,4,5-трифосфат, диацилглицерин, фосфатидная кислота, продукты перекисного окисления липидов мембран. Все эти соединения образуются из липидов плазмалеммы под действием специальных ферментов, активируемых в результате воздействия сигнала на рецепторы. Таким образом, плазматическая мембрана является местом не только рецепции сигналов, но также их усиления и дифференцирования.
Транспорт веществ через плазмалемму
Перенос веществ через мембрану может идти пассивным и активным путем. При пассивном поступлении веществ через мембрану основой переноса является диффузия. Скорость диффузии зависит от толщины мембраны и от растворимости вещества в липидной фазе мембраны. Поэтому неполярные вещества, которые растворяются в липидах (органические и жирные кислоты, эфиры), легче проходят через мембрану. Однако большинство веществ, которые важны для питания клетки и ее метаболизма не могут диффундировать через липидный слой и транспортируются с помощью белков, которые облегчают проникновение воды, ионов, сахаров, аминокислот и других полярных молекул в клетку. В настоящее время показано существование трех типов таких транспортных белков: каналы, переносчики, помпы (рисунок 11).

Рисунок 11 — Три класса транспортных белков:
1 — белковый канал; 2 — переносчик; 3 — помпа
Каналы — это трансмембранные белки, которые действуют как поры. Иногда их называют селективными фильтрами. Транспорт через каналы, как правило, пассивный. Специфичность транспортируемого вещества определяется свойствами поверхности поры. Как правило, через каналы передвигаются ионы. Скорость транспорта зависит от их величины и заряда. Если пора открыта, то вещества проходят быстро. Однако каналы открыты не всегда. Имеется механизм «ворот», который под влиянием внешнего сигнала открывает или закрывает канал. Долгое время представлялась труднообъяснимой высокая проницаемость мембраны (10 мкм/с) для воды — вещества полярного и нерастворимого в липидах. В настоящее время открыты интегральные мембранные белки, представляющие канал через мембрану для проникновения воды — аквапорины. Способность аквапоринов к транспорту воды регулируется процессом фосфорилирования. Было показано, что присоединение и отдача фосфатных групп к определенным аминокислотам аквапоринов ускоряет или тормозит проникновение воды, но не влияет на направление транспорта.
Переносчики — это специфические белки, способные связываться с переносимым веществом. В структуре этих белков имеются группировки, определенным образом ориентированные на наружную или внутреннюю поверхность. В результате изменения конформации белков вещество передается наружу или внутрь. Поскольку для транспорта каждой отдельной молекулы или иона переносчик должен изменить конфигурацию, скорость транспорта вещества в несколько раз меньше, чем происходит перенос через каналы. Показано наличие транспортных белков не только в плазмалемме, но и в тонопласте. Транспорт с помощью переносчиков может быть активным и пассивным. В последнем случае такой транспорт идет по направлению электрохимического потенциала и не требует затрат энергии. Этот тип переноса называется облегченной диффузией. Благодаря переносчикам он идет с большей скоростью, чем обычная диффузия.
Согласно представлениям о работе переносчиков, ион (М) реагирует со своим переносчиком (X) на поверхности мембраны или вблизи нее. Эта первая реакция может включать или обменную адсорбцию, или какое-то химическое взаимодействие. Ни сам переносчик, ни его комплекс с ионом не могут перейти во внешнюю среду. Однако комплекс переносчика с ионом (MX) подвижен в самой мембране и передвигается к ее противоположной стороне. Здесь этот комплекс распадается и высвобождает ион во внутреннюю среду с образованием предшественника переносчика (X´). Этот предшественник переносчика снова передвигается к внешней стороне мембраны, где вновь превращается из предшественника в переносчик, который на поверхности мембраны может соединиться с другим ионом. При введении в среду вещества, способного образовать прочный комплекс с переносчиком, перенос вещества блокируется. Опыты, проведенные на искусственных липидных мембранах, показали, что перенос ионов может проходить под влиянием некоторых антибиотиков, вырабатываемых бактериями и грибами, — ионофоров (рисунок 12). Транспорт с участием переносчиков обладает свойством насыщения, т. е. при увеличении концентрации веществ в окружающем растворе скорость поступлении сначала возрастает, а затем остается постоянной. Это объясняется ограниченным количеством переносчиков.
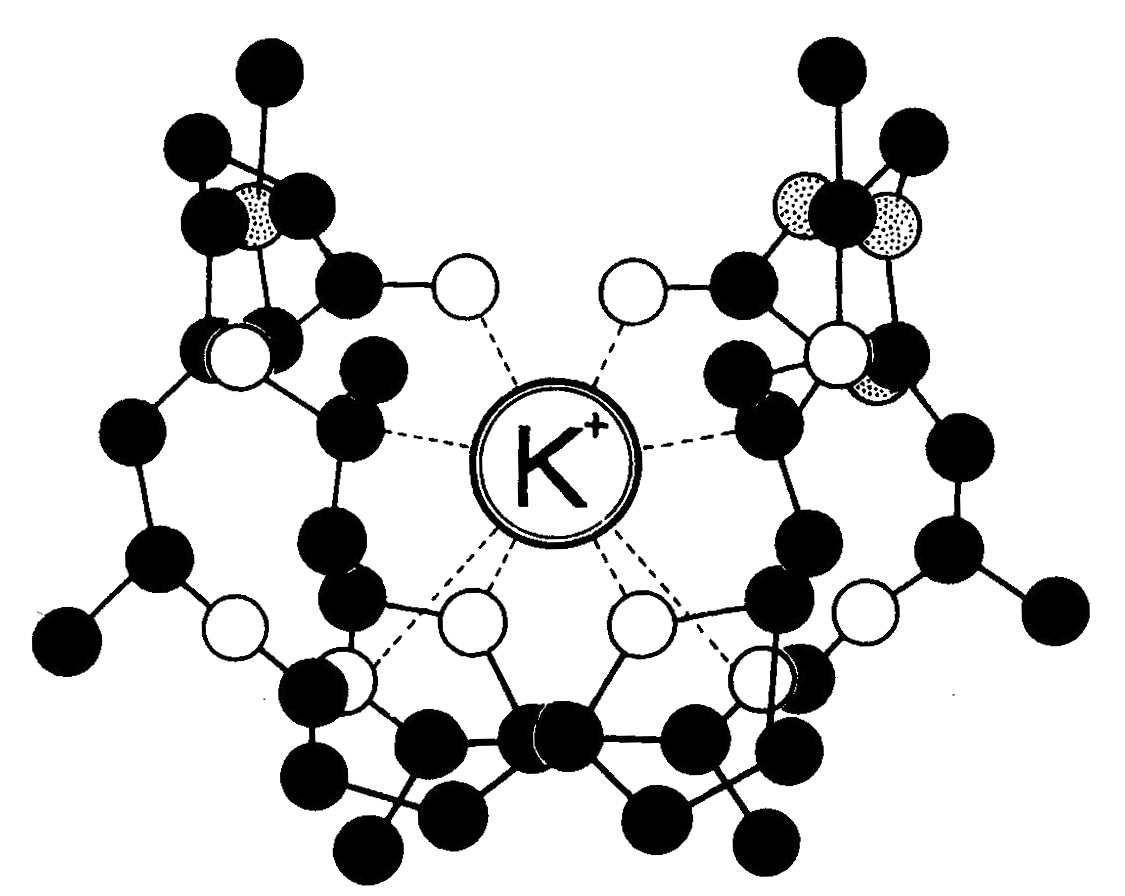
Рисунок 12 — Комплекс ионофор – К+
Переносчики специфичны, т. е. участвуют в переносе только определенных веществ и, тем самым, обеспечивают избирательность поступления. Это не исключает того, что один и тот же переносчик может обеспечивать перенос нескольких ионов. Например, переносчик К+, обладающий специфичностью для этого иона, также переносит Rb+ и Na+, но не транспортирует Сl- или незаряженные молекулы сахарозы. Транспортный белок, специфичный для нейтральных кислот, хорошо переносит аминокислоты глицин, валин, но не аспарагин или лизин. Благодаря разнообразию и специфичности белков осуществляется избирательная их реакция с веществами, находящимися в среде, и, как следствие, их избирательный перенос.
Насосы (помпы) — интегральные транспортные белки, осуществляющие активное поступление ионов. Термин «насос» показывает, что поступление идет с потреблением свободной энергии и против электрохимического градиента. Энергия, используемая для активного поступления ионов, поставляется процессами дыхания и фотосинтеза и в основном аккумулирована в АТФ. Как известно, для использования энергии, заключенной в АТФ, это соединение должно быть гидролизовано по уравнению АТФ + НОН → АДФ + Фн. Ферменты, осуществляющие гидролиз АТФ, называются аденозинтрифосфатазы (АТФазы). В мембранах клеток обнаружены различные АТФазы: К+/Na+–АТФаза; Са2+– АТФаза; Н+–АТФаза. Н+–АТФаза (Н+–насос или водородная помпа) является основным механизмом активного транспорта в клетках растений, грибов и бактерий. Н+– АТФаза функционирует в плазмалемме и обеспечивает выброс протонов из клетки, что приводит к образованию электрохимической разности потенциалов на мембране. Н+–АТФаза переносит протоны в полость вакуоли и в цистерны аппарата Гольджи.
Расчет показывает, что для того, чтобы 1 моль соли диффундировал против градиента концентрации, необходимо затратить около 4600 Дж. Вместе с тем при гидролизе АТФ выделяется 30660 Дж/моль. Следовательно, этой энергии АТФ должно хватить для транспорта нескольких моль соли. Имеются данные, показывающие прямо пропорциональную зависимость, существующую между активностью АТФазы и поступлением ионов. Необходимость молекул АТФ для осуществления переноса подтверждается еще и тем, что ингибиторы, нарушающие аккумуляцию энергии дыхания в АТФ (нарушение сопряжения окисления и фосфорилирования), в частности динитрофенол, тормозят поступление ионов.
Насосы делят на две группы:
1) электрогенные, которые осуществляют активный транспорт иона какого-либо одного заряда только в одном направлении. Этот процесс ведет к накоплению заряда одного типа на одной стороне мембраны;
2) электронейтральные, при которых перенос иона в одном направлении сопровождается перемещением иона такого же знака в противоположном либо перенос двух ионов с одинаковыми по величине, но разными по знаку зарядами в одинаковом направлении.