

Трихомы. Классификация, функции. Значение для диагностики лекарственного сырья
У огромного большинства высших растений клетки эпидермы образуют выросты ? трихомы или волоски (греч. трихос ? волосок). К трихомам относятся самые разнообразные выросты эпидермы. Некоторые из них действительно напоминают формой волоски, другие имеют вид сосочков, бугорков, крючочков, чешуек.
Трихомы бывают железистые и кроющие. В железистых трихомах накапливаются экскреты, поэтому их относят к выделительной системе.
Кроме того, различают волоски одноклеточные и многоклеточные, мертвые и живые.
Мертвые волоски лишены протопласта, полости их заполнены воздухом, вследствие чего они кажутся белыми. Растение густо покрытое мертвыми волосками, имеет седой вид. Такие волоски лучше отражают солнечные лучи и этим уменьшают нагревание и испарение у растения.
Форма волосков очень разнообразна и характерна для того или иного вида растения. Волоски бывают головчатые, звездчатые, крючковидные, чешуйчатые, ветвистые.
Нередко трихомы защищают растения от насекомых. При этом, чем гуще опушено растение, тем реже насекомые посещают его и используют в качестве пищи и для откладки яиц.
От трихомов следует отличать эмергенцы (лат. emergere – выдаваться) – структуры, в образовании которых принимает участие не только эпидерма, но и глубже расположенные ткани. У некоторых растений, малин, роз, образуются эмергенцы, называемые шипами. В образовании шипов у шиповника, например, кроме эпидермы участвуют 2 ниже лежащих слоя. От настоящих колючек (метаморфозов органов) эмергенцы отличаются беспорядочным расположением.
Устичный аппарат. Строение, функции и работа устичного аппарата. Типы устичных аппаратов
Наличие непрерывного слоя кутикулы лишило бы растения возможности какого-либо газообмена со средой, что неизбежно привело бы его к гибели. Поэтому в процессе эволюции возникли специфические структуры ? устьица. Через них и осуществляется сообщение с внешней средой. Там, где нет кутикулы, например, у подводных растений, нет и устьиц. Чем толще кутикула, тем многочисленнее устьица. Через устьица проходит чрезвычайно интенсивная диффузия водяного пара, кислорода и углекислого газа.
Каждое устьице состоит из пары замыкающих клеток и устьичной щели, которая представляет собой межклетник. Замыкающие клетки отличаются от окружающих их обычных эпидермальных клеток своей формой и наличием хлоропластов. Чаще всего замыкающие клетки имеют бобовидную форму. Кроме того, замыкающие клетки обычно имеют более мелкие размеры.
Как правило, замыкающие клетки окружены так называемыми побочными клетками устьиц, отличающимися морфологически от основных клеток эпидермы. Побочные клетки функционально тесно связаны с замыкающими и составляют вместе устьичный аппарат (или устьичный комплекс). Побочные клетки устьиц весьма разнообразны по форме, для них даже разработана специальная классификация, а примитивные высшие растения вообще не имеют побочных клеток.
Раскрывание и закрывание устьиц представляет чрезвычайно важное явление в жизни растений. Полностью механизм работы устьичного аппарата был выявлен совсем недавно, но уже со времен Швенденера известно, что основным фактором здесь является изменение тургора (осмотического давления) внутри замыкающих клеток.
Раскрыванию устьиц, кроме того, способствует неравномерно утолщенные оболочки замыкающих клеток. Внутренние стенки, окаймляющие устьичную щель, более толстые, чем наружные. Поэтому при повышении давления в замыкающих клетках наружные стенки изгибаются сильнее и устьичная щель приоткрывается.
Изменение тургорного давления в замыкающих клетках обусловлено изменением в них концентрации ионов калия. Ионы калия закачиваются в замыкающие клетки против градиента концентрации. На это требуется большое количество энергии, поэтому замыкающие клетки содержат многочисленные митохондрии. Углеводы, необходимые для активной деятельности митохондрий, синтезируются хлоропластами.
При высокой концентрации калия вода всасывается в замыкающие клетки, их объем увеличивается и устьице открывается.
Отток ионов калия и соответственно воды совершается пассивно.
Резервуаром ионов калия служат побочные клетки.
В движении устьиц особое значение имеет также и радиальная ориентация целлюлозных микрофибрилл в оболочках замыкающих клеток. Эти радиальные мицеллы позволяют замыкающим клеткам удлиняться и одновременно не дают им расширяться.
В большинстве случаев устьица в значительно больших количествах расположены на нижней стороне листовых пластинок, чем на верхней. В этом случае устьица не подвержены прямому воздействию солнечных лучей и меньше нагреваются.
Устьица на верхней стороне листа преобладают у травянистых растений, обитающих на сильно нагреваемых каменистых склонах.
И, наконец, у водных растений, таких, как кувшинки, водной лилии, у которых листья расположены на поверхности воды, устьица находятся только на верхней стороне листа.
Таким образом, количество устьиц и их локализация в значительной мере зависит от экологических условий. В среднем на 1мм2 поверхности листа насчитывается 100-300 устьиц.
Строение и работа устьичного аппарата
В состав устьичного аппарата входят две замыкающие клетки (собственно устьица), побочные, или околоустьичные клетки, а также подъустьичная полость.
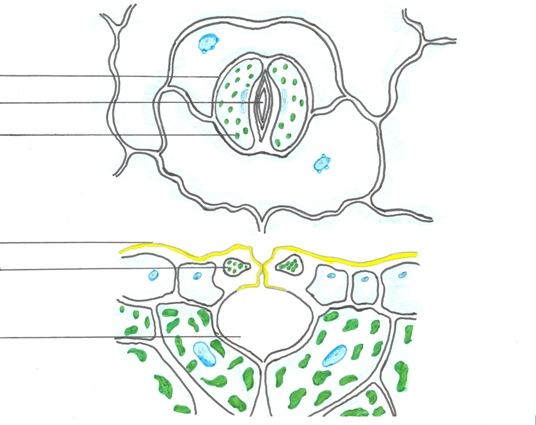
З амыкающие
клетки имеют бобовидную форму, содержат
хлоропласты. Между ними располагается
межклетник – устьичная щель. Устьичная
щель может расширяться и сужаться,
регулируя транспирацию и газообмен.
Стенки замыкающих клеток, ограничивающих
устьичную щель, утолщены сильнее, чем
наружные. Наружные и внутренние стенки
покрыты кутикулой, которая образует
клювовидные выросты над входом в
устьичную щель и на ее противоположном
внутреннем конце. На поперечном срезе
замыкающих клеток видно, что над устьичной
щелью и под ней образуются два расширения.
Наружное расширение называется передним
двориком, а внутреннее – задним двориком.
амыкающие
клетки имеют бобовидную форму, содержат
хлоропласты. Между ними располагается
межклетник – устьичная щель. Устьичная
щель может расширяться и сужаться,
регулируя транспирацию и газообмен.
Стенки замыкающих клеток, ограничивающих
устьичную щель, утолщены сильнее, чем
наружные. Наружные и внутренние стенки
покрыты кутикулой, которая образует
клювовидные выросты над входом в
устьичную щель и на ее противоположном
внутреннем конце. На поперечном срезе
замыкающих клеток видно, что над устьичной
щелью и под ней образуются два расширения.
Наружное расширение называется передним
двориком, а внутреннее – задним двориком.
Побочные клетки устьиц располагаются вокруг замыкающих и, как правило, отличаются по форме от замыкающих клеток. Число и расположение этих клеток разлияаются в зависимости от вида растения.
Устьичная полость – это большой межклетник под устьицем.
Механизм открывания и закрывания устьиц зависит от осмотического давления замыкающих клеток устьиц и клеток их окружающих. Если осмотическое давление в клетках устьиц выше, чем в соседних, то в них начинает поступать вода, и они увеличиваются в размерах. При этом возрастает давление на клеточные стенки, которые начинают растягиваться, и устьица открываются. Осмотическое давление, в свою очередь, регулируется множеством факторов: интенсивностью фотосинтеза в замыкающих клетках устьиц, обводненностью клеток, содержанием осмотически активных веществ.
Наряду с классификациями морфологических
или т опографических
типов устьичного аппарата за последнее
десятилетие появились классификации
онтогенетических их типов. Первая
попытка классифицировать онтогенетическиетипы
была предпринята индийским ботаником
Д. Пантом в 1965 г. Он выделил три крупные
категории устьичных аппаратов в
зависимости от того, происходят ли
клетки, окружающие устьице, из той же
самой материнской клетки, что и устьице,
или из других клеток:
опографических
типов устьичного аппарата за последнее
десятилетие появились классификации
онтогенетических их типов. Первая
попытка классифицировать онтогенетическиетипы
была предпринята индийским ботаником
Д. Пантом в 1965 г. Он выделил три крупные
категории устьичных аппаратов в
зависимости от того, происходят ли
клетки, окружающие устьице, из той же
самой материнской клетки, что и устьице,
или из других клеток:
1) мезогенный (от греч. mesos — средний и genos — происхождение) устьичный аппарат, у которого все побочные клетки или одна кольцеобразная побочная клетка происходит из той же самой материнской клетки, что и замыкающие клетки;
2) мезоперигенный устьичный аппарат, у которого только одна из окружающих клеток образуется таким путем, остальные образуются из других клеток;
3) перигенный (от греч. peri — около,, кругом) устьичный аппарат, у которого ни одна из окружающих клеток не происходит и» той же самой материнской клетки, что и замыкающие клетки.
Таким образом, в основу классификации Панта был положен способ развития устьичного аппарата без учета типов взрослых структур, тогда как классификация морфологических типов строилась исключительно на расположении побочных клеток в зрелом состоянии, при полном игнорировании способа развития устьичного аппарата. Только появление в 1973 г. классификации онтогенетических типов устьичного аппарата, разработанной Е. Фринс-Классенсом и В. Ван-Коттемом, устранило разрыв между морфологической и онтогенетической классификациями.
С эволюционной точки зрения наиболее примитивным среди высших растений является аномоцитный тип устьичного аппарата: он обнаружен у первых их представителей — риниофитов. Устьица у современных мхов и антоцеротовых также обычно аномоцитные, хотя у некоторых мхов устьица окружены кольцом из более или менее дифференцированных клеток