

Шпоры по биофизике
.pdf*31.Особенности межмолекулярных взаимодействий в мембранах.
Бывают взаимодействия: липид липидное, белок липидное, белок белковое. Фосфолипиды являются струтктурообразующими ФАА ФАВ ФВВ
Дисперсионное взаимодействие:
α = ФАВ − ФАА +ФВВ → 0
2
α=0,1;0,2;… есть некоторое расслоение мембраны
–образование доменов. Наиболее резко это взаимодействие отличается тогда, когда молекулы имеют различное количество звеньев в С Н хвостах
–меняется α и на поверхности мембраны возникает домен.
Фазовый переход носит особенный характер. При α >0 он происходит при одной температуре; при
α≠ 0 возможны две или больше точек фазовых
переходов. Эти переходы наблюдаются тогда, когда в состоянии мембраны имеется достаточное количество липидов с ненасыщенными связями. Таким образом липид липидное взаимодействие меняется к изменению фазового перехода.
Липид липидное взаимодействие.
При рассмотрении существенное значение имеет дост во мембранное соотношение между однородными молекулами и между разнородными молекулами. Пусть имеется два типа липидов А и В. Взаимодействие между молекулами А А = ФАА, между В В =ФВВ, между А В =ФАВ.
В зависимости от заданных коэффициентов ФАВ – (ФАА + ФВВ ) / 2.
Если эта разность велика, т. е. взаимодействие между разными молекулами >, че одинаковыми молекулами, то распределение липидов является неравномерным, происходит расслоение мембраны. Основной вклад в межлипидное взаимодействие вносится дисперсионным взаимодействием. Это наиболее общий тип взаимодействия , обусловленный мгновенными нарушениями распределения взаимодействующих молекул. Дипольный момент головки липидов приводит к возникновению диполь дипольного взаимодействия, которое вносит значительный вклад в межлипидное взаимодействие. Существуют липиды, имеющие нескомпенсированный эл. заряд: ион ионные и ион дипольные взаимодействия. Поскольку биологические функции мембран проявляются в водном пространстве, на эти взаимодействия оказывает влияние водное окружение. Молекулы воды чаще всего через водородные связи образуют гидратную оболочку, в часности поляризованная головка всегда окружена молекулярным слоем воды. Это обстоятельство усложняет строгое кванто химическое взаимодействие. Водородная связь является распространенным видом взаимодействий при липид липидном взаимодействии. Одновременно взаимодействие зависит от величины углеводородных хвостов. В зависимости от соотношения насыщенных и ненасыщенных связей меняется характер взаимодействий, влияет на точку фазового перехода. Влияние количества типов молекул на свойства мембраны изучается на синтетических моделях мембраны. Состав естественных мембран точно неизвестен.
Липид белковое взаимодействие.
Является существенным для понимания механизма биологических функций. Взаимодействие белков с липидами делает эту задачу еще более сложной. Л Б взаимод. состоит из : 1) дисперсионные взаимодействия (затухающие),
2)возникновение водородной связи 24 ккал/моль,
3)электростатическое взаимодействие (происходит между полярной головкой и фрагментами белков, являющимися эл. заряженными). Белки находятся в окружении молекул липидов. Это окружение влияет (как экспериментально доказано) на биологические функции белков. В зависимости от липидного окружения, ферментативные белки порой увеличивают активность, порой уменьшают
её. при взаимодействии липидов с белками происходит изменение конфигурации липидов, при котором углеводородные хвосты или раздвигаются, или сдвигаются. При раздвижении – мембрана делается рыхлой и проницаемой. При сближении – сечение уменьшается, мембрана становится плотной, проницаемость уменьшается. Белок липидное взаимодействие:
Особенности классификации – (дисперсионное, водородное, ион ионное – остаются):
1. Белок – монослой; 2. Белок бислой. Эти взаимодействия приводят к тому, что молекула белка принимает определенное положение относительно мембраны. Взаимодействие белок бислой определяет погружение белка в мембрану. Это погружение оказывает влияние на окружающие молекулы липидов – это влияние – до 4 х слоев в латеральной плоскости мембраны. Белки также испытывают влияние липидов. Влияние белков ярко проявляется в поведении ферментов: существуют ферменты, у которых максимальная активность проявляется только в окружении липидов определенного состава. В то же время нет метода определить какой набор липидов как влияет на активность белков. Количество ферментов – несколько тысяч.
У некоторых ферментов активность падает в окружении липидов определенного состава. Но есть факты, когда нет корреляции между микровязкостью и активностью. На активность могут влиять как насыщенные, так и ненасыщенные хвосты.
Белок белковое взаимодействие.
Между белками – дисперсионное взаимодействие, но принято считать, что между молекулами белка преобладает электростатическое взаимодействие. Диполь дипольное взаимодействие между молекулами белков приводи т к относительной ориентации белков относительно друг друга. Белок белковое взаимодействие:
В основном носит дисперсионный электростатический характер, характерно образование водородных связей. Это взаимодействие представляет интерес с точки зрения ферментативного катализа. Агрегации белков приводят приводят к понижению активности. Процесс агрегации зависит от рН: рН>5,5 – образуются межбелковые соединения рН<5,5 – агрегаты разрушаются.
Белок бислойное взаимодействие.
Это взаимодействие является чрезвычайно сложным. Сложность – мембрана не является однородной, поэтому как правило молекулы белка помещают в некоторое усредненное поле, созданное всеми зарядами. имеющимися в бислое. Такой же подход при рассмотрении липид бислойного взаимодействия. Исходя из этих взаимодействий, в идеале, мы должны объяснить свойства мембраны, т.е. возникновение биопотенциалов, проницаемость, избирательные свойства мембраны. Такую задачу решить не удается.
*33.Ионное равновесие на границе раздела фаз. Уравнение Нерета.
Клетки в нормальном биологическом состоянии нах ся в сост ии равновесия, т.е. электр ие потенциалы уравновешены.
μ0 +RT ln c1 +Fz φ1 = μ0 +RT ln c2 + Fzφ2
Это уравнение справедливо, если рассматривать транспорт одного иона. Если ионов много , то
Fz(φ1 φ2) = RT ln(c2 /c1); φ1 φ2 =(RT/Fz) ln(c2 /c1) – Ур ние Нерста. Разность φ м/у межклет средой и цитоплазмоц опр ся как ln относит х концентрации в 2 х средах (в клетке и и вне клетки одного иона). Это Ур ние подтверждается экспериментальными фактами. На перенос ионов накладывается процесс гидратации ионов – возникает гидратная оболочка
– происходит ув ие (эф?)фиктивного радиуса ионов, поэтому меняются условия движения для ионов. На границе раздела фаз происходит в основном гидрофобное воздействие, т.е. гидрофобные части мол л отталкиваются от мол л воды и за счет этого возникает нек упорядочение. Границы раздела фаз могут быть м/у водой и липидами( в основном). Ионы воды разворачиваются противоположным зарядом к мол ле и если взаим вие дост сильно, мол лы воды разделяются на Н+ ОН. Происходит связывание иона водорода и высвобождение гидроксильной группы ОН. Эти процессы исп ся орг мом для расщепления. Если частицы диффундирующего в ва по размеру близки к мол ле воды, то резко меняется вязкость для данной частицы. Гидродинамическое трение зависит от радиуса и вязкости: F=6π r nV. Кроме гидродинамического изменения ионы испытывают диэлектрические силы трения, пропорциональные коэфф ту Fд = kq2 /r3; r средн. радиус объема диффундирующей пов ти. Существует неск ко мех мов транспорта ионов ч/з мембрану: 1. растворение ионов в липидах и дальше обычный мех м диффузии в трансмембранном направлении и переход во 2 ую среду. 2. движение по ионным каналам 3. перенос с участием мол л переносчиков. Основн. движущая сила транспорта электрохим потенциал φ: μ= μ0 +RT ln c нет электр поля. μ0 – стандартный потенциал, кот есть работа, совершаемая молем в ва, растворенным в 1 литре воды при перемещении из данной точки в ∞. Когда появляется эл поле, выражение меняется: μ= μ0 +RT ln c+Fz φ, z валентность,F число Фарадея. Работа по перемещен иона складывается из 2 х частей: по преодолению сил хим взаимодействия и сил взаимодействия иона с эл полем мембраны.
*34.Уравнение Гольдмана.
Выражение описывающее мембранный потенциал: Это ур ние Гольдмана. Оно дает возм ть определить мембранный понциал. При известных св вах иновов. Является одним из немногих ур ний, достаточно точно описывающих значение потенциала.
Зависимость явно нелинейная. P проницаемость для данного иона (хар ка мембраны) Pi= ; γ распределение ионов в области непосредственного примыкания к мембране, h толщина мембраны, u подвижность мол л, z валентность иона. Потенциал Гольдмана справедлив только для установившегося состояния. Для сост я возмущений это ур ние дает невнрные результаты.
Для стац ных состояний, когда конц ция ионов низкая ур ние Гольдмана дает удовлетворительное соответствие с экспериментальными данными.
Для данной конкретной мембраны состояние: для ионов калия примем =1, для Na 0,04, для Cl 0,45:

*35.Блокировка ионного канала.
Пусть входной участок канала имеет при данной концентрации вероятность θ оказаться занятым. Вер ть оказаться свободным (1 θ). Будем считать, что поток ионов, втекающих во вход канала, и обратный поток равновесны.
k1cθ = k2c(1 θ)
k1cθ = k2 c k2 θ; θ = |
k2 c |
= |
|
k2 |
|
k1c + k2 c |
k1 |
+ k2 |
|
|
|
Т.е. вероят ть θ опр ся константами равновесия. Чем выше концентрация, тем больше вероятность того, что потенциальные ямы окажутся занятыми ионами.
Так как электростатические взаимодействия таковы, что в занятое положение не может переместиться новый ион, то данный канал оказывается блокированным. Суммарный поток ионов определяется как сумма отдельных видов ионов, участвующих в движении ч/з данную мембрану.
Метаболические процессы в основном представляются проблемой транспорта. Эта проблема транспорта ионов ч/з мембрану может быть описана в рамках строгих квантовомеханических представлений с учетом таких явлений как туннельный эффект.
Блокировка ионного канала – 2
Блокираторы избирательно перекрывают канал К или Nа. Избирательность достигается за счет геометрии каналов. Канал Nа в сечении 0.3∙0.5 нм, канал К 0.45∙0.45 нм. В каналах происходят сильные ион ионные взаимодействия.
*36.Доннасовское равновесие.
Наблюдается в том случае, когда две среды разделены м/у собой или мембраной, пропускающей достаточно мелкие мол лы и частицы и непроницаемой для нек/т мол л носителей заряда, или сама мембрана обладает фиксированными зарядами. Вокруг фиксированных зарядов образуется слой противоионов, т.е. связанные заряды окружаются ионами противоположного знака и возникает нек/т динамическая с ма, создающая эл. Поле ,причем если связанныезаряды стремятся создать с му распределения зарядов, то тепловое движение стремится разрушить это упорядочение. Это равновесие принято называть доннановским равновесием. Потенциал, возникающий при этом ≈ 5 мв, является сущ ным при толковании
метаболических процессов.
∆ϕ=
При наличии связанных зарядов (плотность):
∆ϕ=
Связанные заряды располагаются вблизи пов ти раздела, они могут иметь неравномерное распределение вдоль поверхности раздела. Следовательно, их влияние на поверхностный потенциал является сложным.
Поверхностный потенциал важен для процессов метаболизма, но измерить его невозможно, т.к. :1) к пов сти мембраны прилипает слой молекул воды, 2)ионы первого слоя, прилегающие к границе раздела, окружены гибридными оболочками.
И для измерения потенциала ξ измеряют скорость движения заряженных мембранных частиц ξ= , где ε электрическое поле, вызывающее движение мембранных частиц, η вязкость, v скорость движения.
*37.Соотношение Уссинга.
Рассмотрим непрерывную теорию. Поток ионов I: I=C∙U∙( dμ/dx), где μ электрохимический потенциал, C концентрация; х координата,U подвижность молекул
μ= |
|
RT |
|
|
|
|
|
|
|
|
|
|
|
||
|
FZ ln C + FZV |
|
|
|
|||||||||||
|
|
|
|
|
|||||||||||
I=CU |
RT |
|
1 |
|
dC |
|
|
dU |
|
||||||
FZ |
C |
|
dx −CUFZ |
dx |
|||||||||||
|
|
|
|
||||||||||||
A |
dC |
+ B |
dϕ |
= CI |
это уравнение |
||||||||||
|
|
dx |
|
|
dx |
|
|
|
|
|
|
||||
элементарно не решается Решением уравнения является величина:
|
|
|
|
|
|
FZϕ |
|
|
|
I = |
UZFϕ |
|
C −C |
e |
kT |
|
где С1, С2 концентрация |
||
|
1 2 |
|
|
|
|
|
|||
h |
|
|
|
|
|
|
|||
|
|
|
FZϕ |
|
|
|
|||
|
|
1 −e kT |
|
|
|||||
|
|
|
|
|
|||||
веществ непосредственно прилегающих к мембране,
h толщина мембраны, U подвижность, Z валентность иона
C1=γ1С0, С0 концентрация раствора в клетке C2=γ2Сi , Сi концентрация в ва вне клетки γ1, γ2 коэффициенты распределения
Полученное уравнение одновременно описывает поток ионов снаружи внутрь и изнутри в межклеточную среду. Решение можно представить в отдельности для потоков ионов каждого направления
|
|
|
|
C |
|
|
C |
e |
ZFϕ |
|||||||
→ |
|
|
|
← |
|
kT |
|
|
|
|||||||
|
I |
= |
1 |
|
|
; I = |
2 |
|
|
|
|
|
|
|||
|
|
|
FZϕ |
|
|
|
|
ZFϕ |
|
|||||||
|
|
1−e |
|
kT |
|
|
|
1 −e |
kT |
|
|
|||||
Отношение двух потоков |
|
|
|
|
|
|
||||||||||
|
|
|
|
→ |
|
|
|
|
|
|
|
|
|
|||
|
I |
= |
|
C1 |
|
уравнение Уссинга, которое |
||||||||||
|
← |
|
|
|
ZFϕ |
|
||||||||||
|
I |
|
|
C2e |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
kT |
|
|
|
|
|
|
|
|
|
|||||
показывает, что процесс движения ионов строго описывается этим соотношением, следовательно, транспорт носит исключительно пассивный характер. Если же наблюдается отклонение от полученного выражения, то имеют место активные процессы транспорта (затраты энергии). Метаболические процессы в клетках происходят с одновременным участием нескольких ионов (например, Na+, K+, Cl) – это основные ионы, определяющие мембранный потенциал. Поэтому полный поток ионов
Iобщ=INa+IK+ICl
В равновесии это уравнение Iобщ=0
Выражение, описывающее мембранный потенциал |
||||||||||
I = |
RT P[K |
|
]+ P[Na |
]+ P[Cl |
] |
|
||||
|
ln |
|
0 |
0 |
i |
|
, где |
|||
ZF |
P[K |
]+ P |
[Na ]+ P |
[Cl |
] |
|||||
|
|
|
i |
i |
i |
i |
0 |
0 |
|
|
Рi проницаемости для данного иона (является характеристикой мембраны, а не среды)
*38.Ионные каналы нервных волокон.
Ионные каналы насквозь пронизывают фосфолипидный слой мембраны. Фосфолипидные молекулы находятся в постоянном движении, что определяет способность мембраны пропускать в ва в цитоплазму и из нее. По
функциональным св вам каналы делятся на 4 группы: насосы,воротные мех мы, индикаторы напряжения, селективные фильтры. Индикаторы напряжения – рецепторы мембран. Представлены белковой молекулой, способной различать биоактивные вещества, взаимодействовать с ними, передавать в клетку информацию о характере биохимического воздействия, способны реагировать на изменение мембранного потенциала. Селективные фильтры – расположены в самом узком месте канала; обеспечивают однонаправленное движение ионов, избирательную проницаемость мембраны, обладают высококаталитической способностью, обеспечивают протекание биореакции как в самой мембране, так и у ее поверхности. Потенциал зависимые каналы бывают открытыми при одних значениях мембранного потенциала, и закрытыми при других. Потенциал независимые каналы стимулируются при воздействии на них химических веществ. Ионные каналы рассматриваются на гигантском аксоне кальмара. Его исследования показали, что вх. и вых. поток явл ся независимым при малых концентрациях. Энергетический профиль для Na+. Профиль описывает много явлений, но он вступает в противоречие с зависимостью потока ионного канала от pH среды. Этот профиль – частный случай ( не зависит от pH среды).Наблюдается, что при pH=5.2 происходит протонирование кислот группами→Блокировка канала объясняется зависимостью потока от степни диссоциации окр. среды. У этого профиля вход иона в точке связывания обеспечивается, но в то же время ион, попавший в точку 2 испытывает трудности с движением вправо, т.е. трудности с преодолением потенциального барьера, высота которого сильно зависит от 1) появления иона в 1 положении, что способно вытолкнуть ион из положения 2 2) конформационного состояния молекул, выстилающего поверхность канала. Геометрический профиль Иногда ионы входят в устье и застревают (канал блокируется).

*39.Натрий калиевый насос.
Транспорт веществ стремится выровнить концентрацию веществ в межклеточной среде и в клетке. Движущая сила – неравномерный заряд или разность концентраций. Но такое равновесие не всегда и не везде. При нормальном функционировании клеток состав и содержание клетки всегда отличается от межклеточной среды (содержание калия в клетке больше, чем в межклеточной среде, а натрия наоборот). При отмирании клетки концентрации выравниваются, значит существует какой то механизм поддерживающий это состояние К и натрия. Это перенос против градиента концентрации. Предполагается, что существуют белки переносчики, образующие комплекс с ионами внутри клетки. После конформационных переходов белок в таком положении, что Na оказывается на наружной стороне мембраны. Здесь под действием ферментов меняется сродство белков переносчиков к Na+. Изменение этого сродства долго изучалось в 50 х годах. Белок переносчик состоит из 2х частей: большого фрагмента и малой субъединицы. Предполагают: связывающий центр Na+ локализован в большом фрагменте, который пронизывает мембрану насквозь. Когда большая субъединица оказывается на внутренней поверхности мембраны, то полностью захватывает 3Na+ (внутренняя полость сферическая – не может вместить больше3Na+ ). В это время малая субъединица своей полостью локализует 2К+. После теплового движения и конформации эти две плоскости совмещаются. Меняется сродство к Na и К. После обмена ионов этой полости возникает новое распределение зарядов, следовательно, конформационное напряжение, которое снимается при возврате большой и малой субъединиц в исходное состояние. Потом опять меняется сродство к ионам.
Молекулы белка сильно ассиметричны: в них есть гидрофобная часть, которая требует большой энергии для соприкосновения с водой и гидрофильная, которая должна оторваться от воды за счет огромной энергии. Конформационные переходы белков не всегда учитывают эту особенность белковых молекул. И механизмы изменения сродства к ионам не понятны.
Этот механизм изучается при быстром замораживании мембран жидким азотом и наблюдают под электронным микроскопом.
*40.Транспорт изотопов (протонов).
Через мембрану транспортируются К, Na, Са, Н. Активный транспорт Са изучен так же хорошо как К и Nа. Существует специальный белок, с которым Са образует комплекс и конформационные переходы переводят Са с одной поверхности мембраны на другую. Вопрос изменения сродства Са и белка переносчика сохраняется. Кв. мех ка не может это объяснить. Транспорт протонов через мембрану может осуществляться 3мя способами:
1.С помощью переносчиков. Переносчики с одной стороны схватывают мембрану, с другой – отпускают (процесс облегченной диффузии). Если концентрация протонов с одной стороны больше, чем с другой, то то этот процесс объясняется частотой соударений.
2.Основан на конфомационном переходе белков, которые протонируются (на какую то их часть садится протон), след но, идет процесс коформ го перехода. В результате фрагмент белка, который был вблизи одной поверхности, оказался на другой поверхности.
3.Транспорт протонов через специальные структуры
СС
О—Н—О—Н… Н—О—Н—О
СС
Такая структура пронизывает всю мембрану. Протон выталкивает водород, а сам встает на его место, и так по цепи. В определенных условиях возникает водородная связь. После этого – поворот Н вокру СО связи. Этот механизм можно обосновать с помощью квантово механических методов, но требуется специальный источник энергии за счет которого будет происходить поворот. Предполагается, что таким источником может быть энергия световых квантов. Расчеты показывают, что форма потенциальных барьеров имеет вид
При случайном соударении протон оказывается в потенциальной яме 1.
Для согласованных поворотов требуется преодолеть более высокий потенциальный барьер. Требуется возбуждение иона, в результате получается др. потенц. барьер
Истолкование переноса протонов важно тем, что транспорт протонов в опред. мере электрически нейтрализует
транспорт электронов. Одновременно протоны являются составляющими многих биохим. реакций. Каждый отдельный этап, каждое конформационное изменение м.б. рассмотрено м дами квантовой химии. Весь процесс в целом объяснить не удается.
*41.Потенциал действия.
При возмущении нервной клетки по ней распространяется потенциал действия. Он возникает под действием порогового или сверхпорогового возмущения.
величина потенциала действия не зависит от величины возмущающего действия.потенциал действия не меняется по мере распределения и не меняется форма импульса. Скорость распространения 1÷100 м/с Феноменологически форму действия можно
объяснить соотношением проницаемостей K, Na, Cl: PK:PNa:PCl=1:0,04:0,045
При возмущении меняется проницаемость мембран для ионов Na и они устремляются внутрь клетки. Потом происходит деполяризация положительный знак относительно среды.При каком то значении потенциала изменяется проницаемость для ионов К→выброс К из клетки. Процесс завершается установлением прежних значений потенциалов.
Электрический сигнал может открывать или закрывать канал Na и K. Канал для ионов Na под действием приложенного поля частиц затыкается механически частицами, другие частицы другого поля открывают. Ионы К не подходят для каналов с Nа. Они индивидуальны. Выделяют соединения, блокирующие избирательно для Na и K. Механизм управления связан с поляризацией полярных молекул, выстилающих канал под действием электрического поля – есть молекулы, у которых под действием электрического поля меняется конформационное состояние. Энергетически более выгодна поляризационная модель.
*42.Дискретное описание ионного транспорта.
При ограничениях: отсутствие гомогенности, наличие нескольких потенциальных барьеров, и т.д. строится дискретная теория транспорта
ионов через мембрану. Она объясняет ряд явлений. При построении теории исходят из след.
приближений. потенц.барьер ч/з мембрану имеет вид вход и выход связаны с процессами гидратации, средн. – селективный карьер. 1
потенц. яма занята, 0 свободна. γ = А∙с∙еxp ( [
Е |
+ |
FϕZ |
])= =с∙ν∙еxp( |
FϕZ |
) A const, с |
RT |
2RT |
|
2RT |
||
концентрация, Е–высота потенциального барьера, ν коэф. переноса, показывает сколько ионов при данной температуре перескакивает ч/з
селективный барьер. |
γ = А∙с∙еxp ( [ |
|||||
|
Е |
− |
FϕZ |
])= =с∙ν∙еxp( |
FϕZ |
)“ ” означает, что |
|
RT |
2RT |
2RT |
|||
|
|
|||||
ионы движутся против поля.Из полученных выражений следует, что завис ть потока ионов от φ, Т нелинейна. Чтобы найти общий поток надо сложить оба потока: γ = γ +γ В общем случае
концентрация ионов внутри мембраны (слева) и справа от мембр. могут быть разными. Но задача упрощается, если предположить что с1=с2 =с
γ = γ +γ = с∙ν∙еxp( FϕZ ) с∙ν∙еxp( FϕZ ) = [sh
2RT 2RT
Fzϕ ] c ∙ν
RT
Следствия: зав ть потока ионов ч/з мембрану опис ся sin ной ф ей, т.е. явл ся существенно нелинейной. Представляет интерес отношение ионов:
r |
|
cν exp(− |
Fzϕ |
) |
|
Fzϕ |
Полученное |
||
|
|
|
|||||||
sj |
= |
|
|
2RT |
= exp(− |
) |
|||
j |
|
cν exp( |
Fzϕ |
|
RT |
||||
|
|
|
) |
|
|
|
|||
|
|
2RT |
|
|
|
||||
выражение дает возм ть сосчитать плотность тока и опр ть ВАХ мембраны.

*43.Токи ворот каналов.
Если предположить, что мембрана является диэлектриком и приложить прямоугольный импульс, регистрируемой длительности, то наиболее существенными будут две составляющие тока: ток разряда
( iC) и ток, связанный с перемещением подвижных воротных частиц (ток ворот).
Ток разряда меняется по экспоненте, направление тока зависит от приложенного напряжения.
Единственныя несимметричная составляющая мембранного тока ток ворот. В нормальных условиях и при приложении прямоугольного импульса:
В аксон кальмара имеется возможность удалить содержимое клетки и залить в нее другой раствор: если убрать Na составляющие, то точка 1 на кривой исчезает.
Составляющая тока мембраны, обусловленная воротным током, явл достаточно малой.
К мембране прикладывают строго симметричные импульсы противоположной полярности. Все симметричные тока будут менять направление в зависимости от знака внешнего поля. Если мембранный ток квантовать и накапливать в памяти ЭВМ, все шумы и симметричные токи взаимно компенсируются, что показывает наличие несимметричного тока, совпадающего по направлению с токами ворот. Чтобы это доказать, нужно удостовериться, что в мембране нет других структур, которые могут внести свой вклад в несимметричную составляющую.
Если принять, что других составляющих нет, то признаками существования токов ворот являются: 1 время релаксации близко к величинам, вытекающим из матем моделей; 2 когда Na канал блокируется Na блокираторами, несимметричная составляющая тока исчезает. Характер экспоненты является близким к тому, что дает матем модель. Величина тока не зависит от темп ры. Кроме аксона кальмара существуют др клетки, где в создание потенциала значительный вклад вносят не только био ионы Na+ и K+, но и Ca2+ в мышечных волокнах потенциал сохраняется приудалении ионов Na+. Поэтому, хотя основные свойства клеток различной природы считаются совершенно одинаковыми, среди нервных клеток, на которые распространяется биологический сигнал, сущ ет 2 типа: 1 в
формировании участвуют ионы Ca2+; 2 не участвуют ионы Ca2+.
*44.Электронно транспортная система.
Электронно транспортная система – это система, которая рассматривает транспорт электронов. Существует несколько механизмов:
1.Туннельный эффект, который истолковывает перенос электронов при сверхнизких t˚, когда движение самих белковых молекул является заторможенным.
Элементарная частица, которая находится в потенциальной яме глубиной Е и имеющая энергию Ео<Е может выбраться из нее (вопреки законам классической физики). Здесь для перехода электрона через барьер должен существовать механизм, понижающий этот барьер.
2.Предполагается, что в молекулах белка есть электронные тропы, которые включают в себя сочетание участков туннелирования, соединенных с участками цепи, имеющих подвижные электроны. Такое сочетание обеспечивает перенос на большие расстояния.
3.С помощью молекул переносчиков, которые имеют структуры типа манжета. В ???????? входит дифференцирующая молекула и в этой оболочке переносится через мембрану. Если концентрация снаружи >> чем внутри, то из за этой разности присоединившаяся молекула отсоединяется от молекулы переносчика. К мембране приложить импульс:
Все симметр. токи будут менять свое направление в зависимости от знака приложенного сигнала, если мембранный ток квантовать и накапливать в памяти
ЭВМ – все импульсы и симметр. токи будут взаимно компенсировать друг друга. Такой эксперимент, проведенный многократно с возрастающей точностью устойчиво показывает наличие несимметр. тока совпадающего по направлению с токами ворот. Чтобы полученный результат убедительно показал, что полученный всплеск тока после накопления связан с токами ворот, мы должны доказать, что в мембране нет других причин, других структур, которые могли бы внести вклад в несимметричные состалвяющие косвенные признаки того, что этот несимм. ток является воротным:
1.Время релаксации тока является близким к величинам, вытекающим из мат. модели. 2.Когда натриевый канал блокируется натриевыми блокаторами, несимметричн. составляющая тока исчезает. Характер экспоненты является близким к тому, который вытекает из мат. модели. Величина тока не зависит от t˚.
*45.Трансформация энергии в первичных процессах фотосинтеза.
При попадании кванта света происходит перенос электрона на более высокий эн ий уровень. Далее существуют следующие воз ти трансформации: в эн ию рассеяния; использ ие эн ии для фотохим процесса; случайная передача.если спектр излучения данной мол лы в данном состоянии, перекрываетсясо спектром др мол лы, то эн ий с 1 ой мол может переходить резонансным методом. Чем больше перекрывание, тем эффективней перенос. Перенос может происходить в рез те столкновения, возможно чисто хим ий перенос В общей схеме можно выделить: 1. Поглощение света ф синтетическими пигментами и миграция энергии к реакционному центру. 2. Под действием происходит первичное разделение зарядов.3. перенос электрона по эл но транспортной цепи. Комплексы светособирающих пигментов молекул, сцепленных слабыми связями, передающие
энергию на фотоактивный центр – ф синтетические единицы –1 ая и 2 ая. Пигменты, входящие в состав 1 ой и 2 ой единиц, разл ся по спектральным св вам. В 1 ой – поглощение света с λ>680нм, 2 ой λ<680нм. Эти ед цы могут возбуж ся селективно. Согласно экспериментам, длинновол ое излучение вызывает окисление цитохрома f (белок), коротковол ое вызывает восстановление его. Т.о. имеется воз ть экспериментально определить процесс вос ния и ок ния. Лазеры короткими и длинными импульсами могут определять время движения возбуждения от ф синт. сис мы 1 к сис ме 2 (порядка 5*10 3с) .
Поглощение света пигментами в собир сис ме идет по 2 каналам.
СС ПБК
СС ПБК –светособир пигмент белковый комплекс, РЦ реакционный центр Движение энергии по 2 м независимым каналам
регулируется за счет конформационного перехода ф синтетических мембранах, при кот меняется топография расположения ПБК ССПБК и, соотв но меняется интенсивность потока энергии к РЦ. Механизм регулирования конформационных состояний зависит от присутствия ионов в среде, pH сис мы. По поводу управления плотностью ионов в среде имеются различные предположения. Основной принцип РЦ: при попадании энергии в акт пигмент происходит отрыв электрона и восстан ие акцептора, далее электрон движется по цепи. Близким по св вам к углероду является кремний, есть предположения, что в реакциях учавствуют белки, где вместо ряда атомов С вкраплены ат Si . если наличие Si будет подтверждено – то может быть построена новая концепция переноса эл ов. При анализе ф биологических процессов трудность связана со временем дислокации эн возб ия (5*10 3с). нужно искать варианты стабилизации возбужденных мол ых сис м, когда будет имется большое время жизни.
*46.Фотопревращения бактериородопеина.
БР сод ся в пурпурных бактериях. Это комплекс белка родопсина и светочувствительного сегмента.
В
этихбактериях происходитпреобразование энергии светового кванта в энергию разности электрохим потенциалов ионов Н+. БР имеет один хромофор (ретиналь). Сходство микробов родопсина и родопсина, кот нах ся в сетчатке глаза человека очевидно: родопсин является почти единственным белком в зрительной сетчатке глаза; хромофором у человека так же является ретиналь , что позволяет использовать бр как некоторую модель зрительной клетки. Ф превращния родопсина сопряжены с ионными и ферментативными процессами, лежащими в основе зрительной рецепции и транс мембранного потенциала. Мол ла бр располагается в строгом порядке в пуппур мембранах в виде диско подобной сплющенной сферы d=0.5мкм и толщина 50 60А.БР располагается в пурп мем строго симметричными группами по 3 мол лы. Каждый тример стабилизирован. Это сост иесохраняется до опред темп ры, кот близка к физиологической. БР состоит из 7 сегментов кот пронизывают мем ну: 3 располагаются верт, 4 имеют наклон к пов ти мембран. Большинство рез ов, кот есть получены из модельных сис м. см. Энергетическая схема первичной стадии фотоцикла.
Энергетическая схема первичной стадии фотоцикла.
Исследуя суспензию уз бактериородопсина(бр), выделенного из пурпурн. мембран, обнаружим, что при комн темп ре, при освещении проходит 6 промежуточных состояний, кот были обнаружены импульсной абсорбционной спектроскопией. Эти сос ия различаются по времени жизни.
Природа ф циклов с первичным этапом превращения бр связана с образ ем стабильного промежуточного сост ия, по своим спектральным хар ам этот неизвестный продукт скорее всего находится в состоянии К610. Методом импульсной лазерной спектроскопии было доказано, что жтот процесс более сложный. Под действием света бр 570 переходит в возб сост ие: бр570...бр570(воз)…J625(нестаб сост 0.5нс)…К610(стаб 5 6нс). Интерессно, что при понижении темп ры до темп ры жидкого азота наблюдается повышение времени жизни К610 в 2 3 р, а при понижении до темп ры жидкого гелия время жизни не меняется. След но, имеется 2 процесса: 1. Зависящие от t 2. Не зависящие (связанные с какими то туннельными механизмами).
Цикл превращения бр сопровождается транспортом Н+ ч/з клеточную мембрану. Перенос протона приводит к возникновению потенц м/у цитоплазмой и межкл средой. Имеются данные о том, что приложенное эл поле влияет на временные соотношения цикла. След но, есть воз ть приложением эл полей влиять на метоболические процессы, проис ие в этой сис ме.
Измененные значения энергии сост ий показывают, что весь цикл превращений обеспечивается за счет эн ии возб ия, получаемой от кванта света.
*47.Проблема цис транс изомерии. Ретиаль.
Глаз – оптическая система, которая фокусирует изображение. На сетчатке изображение перевернутое. Оптическая система глаза выделяет УФ излучение. Обнаружили бактерии, которые обладают пурпурной мембраной, в которой находится белок ретиналь. Эксперименты с пурпурными мембранами позволили объяснить механизм преобразования света в нервное возбуждение. Существуют единственный вид молекул (тела), который воспринимает свет квантами света, при этом ретиналь переходит в другое конформационное состояние. Стерические проблемы: 2 конформационных состояния (1й комплекс с опсином, 2й не способен образовывать комплекс). Комплекс опсин ретиналь возникает только с одной из конформаций ретиналь:
|
СН3 |
Н |
Н |
СН3 |
СН3 |
|
|
|
|
|
|
|
|
||||
|
|
8 9 |
|
13 |
|
|
|
|
6 |
7 |
1011 |
15 |
|
О |
|||
12 |
|
|||||||
|
|
Н СН3 |
СН3 |
14 |
|
|
|
|
|
|
Н |
|
|
||||
|
|
|
|
|
|
|
||
При попадании света возбуждаются π электроны→электрон переходит на разрыхляющую орбиту→высота потенциального барьера м/у 2мя изомерами существенно уменьшается. У теплокровных происходит конформационный переход из цис в трансконформацию. Под действием света с 11го атома происходит поворот. Переход ретиналя в возбужденное состояние непосредственно связан с воздействием кванта света. Дальнейшие процессы многочисленны и сложны. Все они происходят за счет энергии света, как следствие разрушается комплекс. Разрушение комплексов→конформационный переход в структуре опсина, к й завершается переносом протонов и возникновением биопотенциалов. С помощью микроэлектродов измерены потенциалы в палочках и колбочках у животных. В отличие от потенциалов в нервных клетках, пот л, вызванный светом состоит из 2х импульсов: 1й – очень слабый потенциал (ранний биопотенциал), 2й поздний биопотенциал. Его природа отлична от возбуждения в нервных клетках. В клетках зрительной системы пот л покоя зависит от концентрации ионов Na. Существенную роль играет Са, к й, может быть блокирует некоторые каналы.
*48.Ионный канал как динамическая структура.
Кроме диффузии движение ионов происходит по каналам с четким сечением, поэтому для каждого вещ ва р ры сечений соотв ют , что не позволяет идти независимому движению ч/з вещ в.
Существуют понятия: проницаемость мембраны объективное и независимое св во мембраны от внеш. среды. Проводимость определяется окружением, если межклеточной среде нет ионов, то проводимость = 0. Избирательные св ва каналов определяются центральным потенц. барьером, его высота зависит фермент ых процессов или процессов в мембране. Движение ионов ч/з каналы происходит под действием тепловых соударений. Высота избир го барьера может изменится, если на пов ти канала вместо + будет . Если поместить меченные ионы в межклеточное вещество, ч/з движение в мембране можно исследовать и узнать 1.имеется насыщающий хар р потока ионов, 2. Канал обладает блокировочностью. Поток ионов ч/з канал, если известно Е
−[ |
E |
+ |
z F ϕ |
] |
|
kT |
2k T |
||||
I x = A e |
|
, A–постоянная |
величина, связанная с природой мемб. z –кол во электронов, Ix в обратном направлении будет при “ z”.
Предположим, что ионный канал имеет нулевое состояние, тогда энергетический профиль этого
канала в первом приближении будет иметь след вид: первый барьер связан с тем что ион переходит из среды ε1 в др среду с более низким ε2 (происходит дегидротация иона). Но если в один из участков поместить ион, меняется энергетический профиль, т.к. поле ионо огромно (≈107В/м). Последующие ионы будут испытывать влияние 1 го иона. Изменение энер. профиля заключается в следующем: 1. Резко
повышается 1 ый барьер; 2.представление такойформы барьера обусловлено тем, что при такой форме его расчеты дают результат, кот совпадет с эксерим. данными. Формы потенц. барьера при наличии иона в сост ии 10 меняется так, что возникают большие трудности попадания во 2 ой барьер. Если ион перескакивает селективный барьер и занимает 2 ое положение, то 1 ое тоже, как правило, занимается следующим ионом, кот. внедряется из среды. Тогда постоянное поступление 2 го иона легко определяется:
|
-W |
e2 |
|
||
K2 = K1 e |
2RT |
W = |
|
, где e заряд |
|
rε |
|||||
|
|
|
|
||
иона, r раст е м/у потенц. ямами, ε диэл. постоянная.
Высота селективного барьера изменяется, т.к. белки, выстилающие канал, мембраны, имеют очень короткое время жизни. Они могут совершать конформационные переходы под действием теплового движения. Существует гипотеза, кот пытается объяснить избирательные св ва каналов мембраны автоматическим изменением высоты селективного барьера из за попадания иона: ион попадая в 1 ую пот. яму под действием куллоновского взаимодействия вызывает такое изменение селективн. барьеров, что высота барьера понижается и ион проходит. Других надежных мех ов, кроме конф переходов белков, выстилающих пов ть канала, не обнаружено.

*49.Физика процессов рецепции. |
*51.Физические поля организма человека. |
Клетка, как часть сложной системы, выживает |
Процессы, происходящие в организме человека, |
только при условии обмена информацией с |
сопровождаются возникновением, изменением |
другими клетками, межклеточным пространством и |
различных полей. 1. Акустическое поле процессы |
окружающей средой. Информация поступает в |
жизнедеятельности, работы сердца, легких, |
клетку и оценивается системой хеморецепсий, |
движение крови и т.д. сопровождается |
которые обеспечивают функционирование самой |
возникновением волновых процессов и человеческ. |
клетки. Большинство сигналов имеет химическую |
организм непрерывно излучает акустич. волны в |
природу, передача сигнала считается с переносом |
состоянии функционирования. Но когда меняется |
молекул носителей информации. Молекула |
волновое сопротивленияе, происходит сильное |
должна пройти расстояние, чтобы доставить сигнал |
отражение. Коэф. отражения на границе воздух |
рецептору, после чего происходит распознавание |
ткань =1 акуст. волны излуч. ослаблено. Ухо |
сигнала (геометрически распознавание объясняется |
согласовано с окружающей средой по волновому |
возможностью взаимодействия только с мол ми |
сопротивлению, след но хорошо воспринимает |
определ ого строения). Если мол ла соответсвует |
волны. 2. Э/м колебания – движение любых |
требованиям, то образуется комплекс, после чего |
связанных з дов , ионов , сопровождается |
вырабатывается сигнал, вызывающий развитие |
возникновением пост. эл., пост. магн. и |
внутриклеточных процессов→ изменения. Стадии: |
переменных э/м полей. Поэтому излучается э/м |
доставка, связывание, узнавание, преобразование |
поле, кот. обусловлено внутр. физиологическими |
сигнала, функциональное изменение, системные |
процессами: до 103Гц – сердце,109 1010Гц – не |
процессы. Сигнальные мол лы воспринимаются |
известно, амплитуды – очень малы. Постоянное эл |
рецепторами, они вырабатываются внутри |
ое поле обусловлено имеющимися, в частности, на |
организма или попадают из окруж среды. |
пов ти тела эл. зарядами. 3.Человек ис точник |
Преодоление расстояний сигнальными мол ми |
теплового излучения, т.к. физиологическая |
достигается за счет конверсии воздуха, воды, |
температура достаточно высокая, согласно E=σT4 |
потока крови и т.д. Время движения сигнальной |
max излучения на λ=10мкм. 4. В организме |
мол лы: |
происходит множество ядерных реакций, сл но |
τ= B2/Dif(A/B), где B² радиус сферы, где находится |
радиационное поле. 5. Человек испускает свет в |
рецептор. |
видимом спектре. Если рассчитать то свет, |
*50.Уравнение Блоха. |
излучаемый пов ю тела на 2 порядка больше |
интенс ти теплового изл. О экстрасенсе: 1. Физ фак |
|
Уравнениями Блоха является система сдвоенных |
р должен проникать ч/з ткань орган ма в |
дифференциальных уравнений, которая |
окружающее пространство и распространяться. 2. |
используется для описания поведения вектора |
Экстрасенс должен обладать св ом воспринимать |
намагниченности в любых условиях. |
этот фак р. 3. Также он должен уметь распознавать |
|
и идентифицировать этот фак р и ставить диагноз. |
|
4. Экст с должен излучать фактор, кот должен |
|
воздействовать на пораженную ткань. |
Правильное интегрирование, уравнения Блоха дает X', Y', и Z составляющие намагниченности, как функции от времени.
*52.Рентгеновская компьютерная томография. Уравнение Радоно.
Исследования внутренней структуры объектов с помощью рентгеновского излучения широко распространены и хорошо известны. Ослабление рентгеновского излучения вдоль луча, соединяющего источник и приемник, является интегральной характеристикой плотности исследуемого объекта.
С математической точки зрения речь идет о задаче восстановления функции по ее интегральным значениям вдоль некоторого семейства лучей. Различные лучи соответствуют различным (относительно объекта) положениям источника и приемника излучения. Такая модель является простейшей, но во многих случаях хорошо отражает реальную ситуацию и подтверждается исследованиям реальных тестовых объектов. Плотность реальных объектов является функцией трех пространственных координат.
Однако в классической компьютерной томографии трехмерный объект представляют в виде набора тонких срезов. Внутри каждого среза плотность считают функцией только двух переменных. При исследовании фиксированного среза систему источник приемник устраивают таким образом, что регистрируются данные только по лучам, лежащим в тонком слое относительно центральной плоскости среза. Таким образом приходят к задаче восстановления функции двух переменных по ее интегральным значениям вдоль некоторого семейства лучей.
Типичные схемы сканирования представлены на рисунках 2 и 3. На рисунке 2 представлена параллельная схема сканирования, обычно используемая в теоретических исследованиях, на рисунке 3 представлена веерная схема, чаще встречающаяся в реальных томографах.
Для регистрации используется линейка детекторов, различные положения источника относительно объекта обеспечиваются вращением системы регистрации или объекта. В непрерывном случае приведенные схемы эквивалентны, однако в реальных ситуациях применение каждой из схем имеет свои особенности, связанные с тем, что число приемников и число положений источника конечно.
В 1917 году Радоном написаны уравнения дающие возможность получить послойное распределение изменения пронизыв х агентов в толще тела.
Как любой матем. аппарат он не связан ни с природой агента, ни объекта исслед я , то есть исп ся и в ЯМР, УЗ …
Пусть есть рентгеновское излучение с интенсивностью в точке

B
I(x, y, z ) = ∫ I0 eφ(x , y , z ) dxdydz
A
φ– коэф т затух я опред й затухание вещ ва
вточке с корд ми (x,y,z)
Эта задача решена Радоном и лежит в основе всех методов.
|
1 |
2 |
|
d |
|
φ(x, y, z ) = |
∫π |
H |
P (r, θ ) d θ |
||
|
π |
0 |
|
dr |
|
Предполагается что объект исслед я просвеч ся многократно, т.к. в единичном случае не хватает числа уравнений.
*53.Флуктуации проводимости мембран.
В мембране сущ ют каналы,к/т насквозь пронизывают весь ее фотолипидный слой. Через каналы движутся ионы K+ Na+ .
На наружный пов ти притягиваются ионы K+,что приводит к накоплению «+» заряда снаружи мембраны, а с другой стороны накапливаются анионы с отриц. Зарядом Появляется разность пот лов, к/т приводит к
открытию калиевых каналов. Ионы калия начинают диффундировать в цитоплазму ч/з мембрану и постепенно уменьшать разность пот лов.
С накоплением калия с внутренней стороны мембраны начинают открываться Са каналы и Nа каналы
Nа каналов больше, чем Са каналов в 20 раз. Потом К+ каналы закрываются(когда разность пот лов уже изменит свой знак), но Са и Nа каналы продолжают функционировать. Разность пот лов опять же увеличивается до тех пор, пока не закроются Са и Nа каналы. Этот момент называют 'активацией'. Вследствие этих механизмов разности пот лов внутри и снаружи мембраны колеблятся и возникает явление, наз мое ' флуктуация проводимости мембран '. За счет этого мембрана активно обменивается информацией с окружающей средой, соседними клетками, межклеточной жидкостью.
Di размерности про ва, А размер сигнальной мол
лы.
Если про во одномерное, то τ= B2/D(1)(1 (A/B)2). При А<<В А/В пренебрегают.
Такое пространство у аксонов кальмара, нервных
клеток чел ка. Если двумерное про во: τ=В2/ D(2)ln(В/А), если трехмерное про во: τ=В2/3АD(3).
Формулы справедливы для расчета времени движения у других препаратов. Предполагают, что в с ме нет вещ в , взаимодействующих с сигнальными рецепторами, и что все реагирующие элементы одновалентны. Комплекс сигнальной мол лы не зависит от остальных и приходит в состояние равновесия по кинетическим принципам Li=Fi (свобод)+ ΣBj (компл), L сигнальные мол лы (леганды). Если L двухвалентны, то добавляют коэф нты сродства: Кij=Bij(число компл)/Fiγj(рецепторы).
*54.Шумы производимые каналами.
Шумы ионных каналов. Каждый из ионных каналов мембраны, обладающих теми или иными воротными механизмами, даже при постоянстве МП и химического состава среды не находится все время в каком то одном положении; он то открывается, то закрывается. Эти переходы от закрытого состояния к открытому и обратно осуществляются по закону случая и практически мгновенно, что обусловливает прямоугольную форму одиночного толчка тока в канале. Время открытого состояния называют "временем жизни канала". Хаотическое открытие и закрытие каналов, порождающее перемещения ионов, создает электрический шум. Для потенциалозависимых каналов соответствующее электрическое поле является фактором резко увеличивающим вероятность открытого состояния каналов, что и создает эффект увеличения ионной проницаемости. Для исследования динамики отдельных каналов определенного вида исследуют слабый электрический шум, создаваемый этими каналами в каком то небольшом участке мембраны в условиях клампа МП и химической блокады прочих каналов. Например, в мембране гигантского аксона кальмара так изучили шумы отдельно натриевых и калиевых каналов.
Анализ этих шумов позволил рассчитать плотность расположения каналов в мембране и проводимость одиночного канала.
В гигантском аксоне кальмара плотность натриевых каналов оказалась равной 300/1 мкм2, проводимость канала 4 рмО. В перехвате Ранвье амфибий плотность натриевых каналов 2000/1 мкм2, а средняя проводимость канала равна 8 рмО.
*56.Распределение импульса вдоль волокна.
Импульсы вдоль волокна распределяются в зависимости от диаметра нервного волокна. По скорости делятся на три группы:1) толстые миелиновые волокна – ск ть >14 м/с, толщина волокна 12 120 мкм; 2) тонкие миелиновые волокна –ск ть 3 14 м/с, толщина 1 3 мкм; 3) безмиелиновые волокна 0,6 0,3 м/с, 0,4 1,2 мкм. Синопс специальное место контакта двух возбужденных стр р , предназначенное для передачи возбуждения. Нервные окончания образуют утолщения, покрытые пресиноптичиской мембраной; в пресиноптическом окончании имеется везикулы (пузырьки) с медиатором, а также большое количество Са каналов, между нервными окончаниями и мышечным волокном имеется синоптическая щель. Часть мембраны мышечного волокна, кот граничит с нервным волокном носит название постсиноптической мембраны. Синопсы способны односторонне проводить возбуждение.
*55.Вариационный метод Ритца.
Пусть имеется набор линейно независимых
функций  , которые мы будем использовать при построении пробной волновой функции Ф:
, которые мы будем использовать при построении пробной волновой функции Ф:
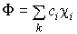 (IV.30).
(IV.30).
Подстановка в (IV.30) функционала энергии (IV.24) дает:
(IV.31).
Поскольку функции 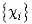 фиксированы, то экстремум функционала энергии достигается при условии:
фиксированы, то экстремум функционала энергии достигается при условии:
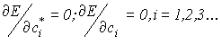 (IV. 32).
(IV. 32).
Выполняя дифференцирование (IV.31), получим систему уравнений:
(IV.33).
Второе равенство в (IV.33) есть комплексно сопряженное от первого, поэтому достаточно рассмотреть одно из них, например первое. Это есть система линейных алгебраических уравнений и она имеет решения, если
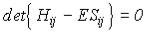 (IV.34).
(IV.34).
Решая уравнение (IV.34), мы можем получить N
корней 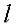 , подстановка которых в (IV.33) дает
, подстановка которых в (IV.33) дает
нам наборы коэффициентов 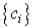 , т.е. пробные
, т.е. пробные
функции 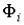 , отвечающие стационарным точкам функционала энергии.
, отвечающие стационарным точкам функционала энергии.
Изложенный выше вариант метода называется вариационным методом Ритца. Он имеет одну замечательную особенность: приближенные
значения энергии 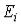 , полученные из уравнения (IV.34) являются оценками сверху для соответствующих собственных значений уравнения Шредингера, т.е. методом Ритца мы можем изучать не только основное, но и возбужденные состояния квантово механической системы.
, полученные из уравнения (IV.34) являются оценками сверху для соответствующих собственных значений уравнения Шредингера, т.е. методом Ритца мы можем изучать не только основное, но и возбужденные состояния квантово механической системы.

*58.Экспериментальные методы исследования биопотенциалов.
Особенности формирования биопотенциала в зрительных клетках.Осн. особ ть в том, что в этих клетках цитоплазма имеет «+» потенциал относительно межклеточной среды, т.е. полярность потенциалов клетки яв ся обратной. Это связано с тем, что в ур и Гольдмана проницаемость для560 ионов Na яв ся преобладающей по сравнению с другими ионами. Концентрация Na в клетке превышает конц ю Na в межклеточной среде. Ур е Гольдмана корректно описывает формир е пот ла в зав ти от конц ии тех или иных ионов. Процесс формирования биопотенциалов яв ся близким к рассмотренному, хотя струк ра биоптенциалов сложнее. Сущ ет ранний потенциал и поздний потенциал. Формир е позднего пот ла после сверхпорогового возбуждения м.б. объяснено конформационными изменениями, происходящими в мембране, а раннего пот ла – с процессами, происх ми в каналах. После действия фотона начинается формир е быстрого потенциала (БРП). Вел на его не превышает 5 мВ. Можно предположить, что БРП в колбочках возникает за счёт движ я связ. зарядов. Оно м.б. обнаружено таким же методом, каким обнаруживаются токи затворов каналов, по кот. двиг ся ионы. Возникает ток, кот. выдел ся методом длительных накоплений и с пом. синхронных детекторов.
Особенности формирования биопот ла в клетках сердечной системы. Снятие кардиопот ла – самый распространённый диагностич. метод. Отличие от формир я пот лов в нервн. клетках в том, что длит сть ПД кардиомицита во много раз >, чем длит сть ПД в нервн. клетках (~1мс), кардиомицита ~ 350мс. Соотнош е конц и ионов Na и K в сердечн. клетках близко к содерж ю в нерв. клетках. При возбуждении клетки, когда наступ. фаза реполяризации, происх. Изменение проницаемости мембран для ионов Ca. В состоянии покоя пот л покоя имеет традиционную полярность (« » в цитоплазме). Движ е ионов внутрь поддерживает к. то время неизменным ПД. После происх. блокировка ионных каналов, кот. селективно пропускают ионы Ca. Пот л падает. Эти каналы имеют электрич. механизм регулирования.
*59.Перенос протонов в фотосинтезе.
Бакрадопсин(БР) 0640
N520
570
М412 (13цис )
L550
М412 – важнейшее состояние в цепи превращений. Оно хар ся 2 факторами: 1)полностью исчезает спектр цис конформации, 2)в это время начинается распад этого состояния, возникают свободные протоны. Эти протоны возникают на другой стороне мембраны. Это определяется по изменению биопотенциалов и по нек. хим. процессам. Это устан ся по косв. признакам.
Переход L550 M412 превышает время деградации энергии. Все процессы происх. За счёт внутр. энергии и не требуют дополн. её источников. Перенос протонов ч/з толщу мембраны осущ ся вдоль цепи аминокислотных остатков за счёт протонирования и депротонирования. Если к этим системам приложить эл. поле, то процесс транспорта протонов меняет скорость. Влияние тем ры на скорость формирования биопотенциалов нелинейное. Сущ ет перелом при тем ре жидк. N, кот. Даёт возм сть предположить, что на процесс транспорта протонов оказывают влияние конформационные переходы в опсине. Интенс ть света завис. от тем ры. Легко измерить процесс депротонирования, заменяя протоны на дейтерий. Все мех мы транспорта протонов реализ ся в мембранах пурпурных бактерий..
*61.Общие характеристики рецепции.
Существование любых биосистем возможно только в условиях обмена информацией, командами, сигналами между отдельными ячейками этой системы и с окружающей средой. В высокоорганизованных системах каждая клетка является автономной системой, с другой стороны она находится в состоянии непрерывного обмена с окружающей средой. В то же время выживание этих систем невозможно без адаптации к среде обитания, которая невозможна без непрерывного обмена с окружающей средой. Большинство факторов говорит о том, что передача сигналов в организме между системами связанос перемещением “сигнальных” мол л. Передача информации между системами в частности носит химич характер. “Сигнальные” мол лы могут перемещаться на расстояния от нескольких мм до нескольких км. В организме “сигнальные” мол лы перемещаются с помощью кровотока, течением лимфожидкости и другими макроспособами. При непосредственной близости от рецептора движение происходит по принципу диффузии. После сближения до расстояния химич связи, процесс диффузии сближает сигнальную мол лу на расстояние взаимодействия с особыми структурами мембран рецепторов. Происходит процесс узнавания –процесс взаимодействия сигнальной мол лы и рецептора. Без взаимодействия невозможен узнавания . Процесс узнавания является триплетом диффузии. После узнавания происходит преобразование этого взаимодействеия (внутриклеточ процесс). Затем следуют, видимо, физиологические изменения.
*60.Стационарные точки. Особенности устойчивости биосистем.
Состояние сист.,наход ся в равновесии в неживой природе опр ся как состояние,кот. Возвращается в исходное сост е после прекращения возбуждения.В биол.сист. такой возврат в точное исходное сост е невозможен,т.к. собств.биол.сист. наход ся в сост и непрерывного и необратимого развития.
Стационарное сост е наз ся устойчвым,если при возмущении сист. Уходит не слишком далеко.Для опр я устойчивости сист. Применяют подход:при выводе из сост я сист.уходит достаточно далеко и быстро,такая сист. наз ся сист. наход ся в неустойчивом стационарном сост и.Пусть сист описывается единств.уравнением,в некоторой
точке X 0 имеется стац.сост е
f( X 0 ) = 0 .Допустим функция в этой точке
x=x0
получает приращение X = X 0 +δ будем
считать δ достаточно малой величиной,кот. Не должна охватывать какой то обл ти,содержащей резкие изменения ф ий,описывающей сост е сист. ассмотрим поведение этой сист:
df ( X 0 |
+δ) |
= |
f ( X 0 ) + |
df (x) |
* |
δ |
+ |
d 2 |
f ( |
dt |
|
dt |
1! |
dt 2 |
|||||
|
|
|
|
|
|||||
df (δ) = a1 *δ + a2 *δ 2 +... вторым dt
членом ряда м/о
пренебречь,т.к. δ 2 << δ : dδδ = a * dt , δ
приращение аргумента в т. стационара; a производная в этой
точке. x = x0 * ea*t ,пусть
C = C0 * ea*t Сист. будет наход ся в
устойчивом сост и,если полученное выражение будет убывать по экспоненте=>производная функции,описывающей сост е сист.,будет отрицат.=>с течением времени сист. будут отклоняться от первоначального сост я достаточно медленно и удаление от этой сист. будет замедляться.Если 1 ая производная от функции по времени положит. В этой точке x,то сист. будет удаляться от этой сист. достаточно быстро и если не будет др. особых точек,сист. уйдет в бесконечность.Сист. неустойчива=>она распадется или приходит в устойчивое сост е.Т.образом,чтобы определить устойчивость стационарного сост я,необходимо опр ть производную в точках прилегающих достаточно близко к стационарному сост ю.Условие=0 содержит в себе еще одно упрощение,оно предполагает,что во всем объеме просранства переменные величины явл ся установившимися,но реально в биол. сист. этого не происходит. В клетке происходят сложн.процессы диффузии.Поэтому уравнение должно приобретать
вид: |
dc |
i |
* f |
|
(c , c |
|
,...с |
|
) = D * |
d 2 c |
i |
|
i |
2 |
n |
|
|||||||
|
|
|
|
||||||||
|
dt |
1 |
|
|
dt 2 |
|
|||||
|
|
|
|
|
|
|
|
||||
,D коэф т диффузии,t обобщенное пространство.

*62.Характеристика начальных стадий |
реакция присоединения, например образование |
*63.Первичный акт фотопревращения радопсина. |
фотобиологических процессов. |
димеров: А + hv * А* + А * (А*...А) * А А + hv фл. |
В настоящее время можно считать полностью |
При поглощении кванта света один из |
|
доказанным, что начальный этап фотоцикла и |
электронов, находящийся на нижнем |
|
образование бато продукта происходит в |
энергетическом уровне на связывающей орбитали, |
|
результате цис транс фото изомеризации ретиналя. |
переходит на верхний энергетический уровень, |
|
Однако доказательство этого факта потребовало не |
заполняя нижнюю разрыхляющую орбиталь и |
|
только больших экспериментальных исследований. |
переводя тем самым молекулу в возбужденное |
|
Необходимо было также достичь теоретического |
электронное состояние (синглетное S1 или |
|
понимания механизмов быстрой |
триплетное Т). Существуют различные пути транс |
|
фотоизомеризации ретиналя в плотной белковой |
формации энергии возбуждения, включающие |
|
среде при участии и других молекулярных групп, со |
внутримолекулярную конверсию, |
|
скоростями, превышающими таковые в модельных |
безызлучательную передачу энергии возбуждения |
|
растворах с малой вязкостью. Мы остановимся |
другой молекуле (миграция энергии) и |
|
кратко на развитии этого направления, где имеется |
использование энергии возбуждения в каких либо |
|
много общего с соответствующими работами в |
фотохимических реакциях: |
|
области фотосинтеза и фотопревращений |
|
|
бактериородопсина. |
|
|
Цис транс фотоизомеризация. Простое |
|
|
сравнение химических конфигураций родопсина |
|
|
(11 цис) и изородопсина (9 цис) показывает, что их |
|
|
взаимопревращение через единственный общий |
|
|
интермедиат батородопсин возможно, если этот |
|
|
процесс включает цис транс фотоизомеризацию. |
|
|
Иными словами, в батородопсине ретиналь должен |
|
|
находиться в транс конфигурации, что |
где кф, км, к — константы скоростей |
|
подтверждено в настоящее |
|
|
|
соответствующих процессов, причем к пред |
|
время большим количеством работ. Если |
ставляет собой суммарную вероятность |
|
исходить из 11 цис , 12 в цис исходной формы |
внутримолекулярной дезактивации возбужденного |
|
ретиналя, то начальная стадия фотолиза родопсина |
состояния [см. (Х.2.8)]: к = р + q + г где р, q, r — |
|
включает следующие изменения конформации |
частоты переходов в единицу времени с |
|
ретиналя: 11 цис, 12 в цис — 11 транс, 12 в цис — |
синглетного возбужденного уровня Si на основной с |
|
11 транс, 12 в транс. |
испусканием света флуоресценции р, тепловой |
|
|
дезактивации состояния Si(q), конверсии Si * Г(г). |
|
Роль протона. Поглощение света и изменение |
Реакционная способность возбужденной |
|
электронного состояния должно привести к |
|
соответствующим молекулярным перестройкам в |
|
светом молекулы определяется рядом факторов. |
|
хромофоре и его ближайшем белковом окружении. |
Она зависит от положения возбужденного |
|
Цис транс изомеризация ретиналя может быть не |
энергетического электронного уровня, что |
|
единственной из таких фотоиндуцированных |
обеспечивает преодоление энергетического |
|
перестроек. Эти вопросы изучали методами |
барьера (рис. XXVII. 1). Большое значение имеет |
|
низкотемпературной спектроскопии, с помощью |
время жизни т возбужденного состояния, в течение |
|
которых исследовали фотопереход родопсин * |
которого сохраняется избыток энергии в молекуле. |
|
батородопсин. Применение абсорбционного флеш |
Во многих фотохимических процессах реализуется |
|
фотолиза с пикосекундным лазерным |
высокая реакционная способность триплетного |
|
возбуждением позволило измерить скорость этой |
состояния. Она обусловлена его относительно |
|
реакции в широкой области температур. Оказалось, |
большим временем жизни (тт >> tsi), а также |
|
что при комнатной температуре время образования |
бирадикальными свойствами. В триплетном |
|
батородопсина < 6 пс, а при 4 К оно составляет 36 |
состоянии молекула обладает двумя |
|
пс (Рентцепис). Кривая зависимости константы |
неспаренными электронами, приобретая тем |
|
скорости от температуры носит двухфазный |
самым свойства анион радикала на время |
|
характер и, кроме того, обнаруживает явно |
существования возбуждения. Известно, что в |
|
выраженный изотопный эффект уменьшения (в 7 |
реакционноспособных анион радикалах, |
|
раз) скорости реакции при дейтерировании (рис. |
полученных путем восстановления нейтральных |
|
XXIX. 17). |
молекул, присоединенный электрон занимает |
|
|
нижнюю вакантную разрыхляющую орбиталь, но не |
|
Фемтосекундные изменения. Эксперименты, |
может понизить свой энергетический уровень и |
|
выполненные с применением фем тосекундной |
перейти в систему связывающих орбита лей, |
|
техники (Матис, Шенк), позволили |
поскольку последние полностью заполнены в |
|
зарегистрировать изменения поглощения |
нейтральной молекуле. |
|
родопсина в области 480 580 нм на временах до |
Собственно фотохимические превращения |
|
500 фс. Оказалось, что выцветание полосы |
|
поглощения родопсина при 500 нм и образование |
|
проходят, как правило, в виде одно квантовых |
|
первичного фотопродукта полностью |
фотохимических процессов. Исключение |
|
заканчиваются в течение 200 300 фс. Это намного |
представляют условия мощного светового |
|
короче, чем время обычной электронно |
лазерного возбуждения, когда за время |
|
колебательной релаксации (10 11 10~12 с). |
возбужденного состояния молекула успевает |
|
|
поглотить второй квант, так что становится |
|
|
возможным двухфотонное возбуждение одной |
|
|
молекулы и переход ее на верхние возбужденные |
|
|
уровни. |
|
|
Как правило, диффузионные процессы в |
|
|
конденсированных средах, которыми являются |
|
|
биологические мембраны, не успевают обеспечить |
|
|
высокую частоту встреч реагентов за время жизни |
|
|
Si состояния. Это затруднение, однако, несуще |
|
|
ственно для триплетного состояния. Оно также |
|
|
облегчается, если молекула образует комплекс со |
|
|
вторым реагентом в основном или возбужденном |
|
|
состоянии — эксиплекс. В этом случае возможна |
|
|