Образовавшаяся в эритроците угольная кислота быстро
диссоциирует. Часть ионов нсо3- диффундируют в плазму, связывая там ионы натрия, в эритроцит же поступают в порядке сохранения ионного равновесия ионы хлора. Ион н' образует комплекс с восстановленным гемоглобином, так как восстановленный гемоглобин обладает более слабыми кислотными свойствами, чем оксигемоглобин и может присоединять дополнительное количество ионов н'.
Кроме того, дезоксигемоглобин за счет уменьшения протонного сродства легче образует карбаминовые соединения, связывая при этом еще около 5-15% переносимого кровью СО2.
С гемоглобином СО2 связывается через аминогруппы белкового компонента молекулы.

Гемоглобин, связанный с СО2, называется карбогемоглобин.
В легочных капиллярах происходит высвобождение части СО2, который диффундирует в альвеолярный газ (рис. 14). Этому способствует более низкое, чем в плазме, альвеолярное Рсо, и усиление кислотных свойств гемоглобина при его оксигенации. Ионы нсо3- поступают из плазмы в эритроцит, а ионы С! - в обратном направлении.

NаНСОз - Na. + н со; !
плазма СГ 1 92

карбоангидраза C02+H20J.H2C03+-H•+ НСОз
НЬ02 + н• +- ННЬ+О2
'------ СО2+ННЬ +- ННЬСО2
--

Глава 111
МЕХАНИЗМЫ РЕГУЛЯЦИИ ДЫХАНИЯ
Организм осуществляет тонкое регулирование содержания кислорода и углекислого газа в крови: их содержание остается относительно постоянным, несмотря на колебания количества доступного кислорода и потребности в нем.
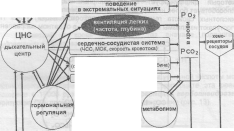
Функциональная система поддержания газового гомеостаза включает многие исполнительные структуры, такие как дыхательная система, система крови, сердечно-сосудистая система, выделительная система (почки, кожа), ВНД (рис. 15). Характерной чертой функциональной системы поддержания газового гомеостаза является конечный результат, включающий несколько взаимосвязанных
параметров - Рсо-, Ро2, рН.
состав воздуха, t? физическая нагрузка эмоционаnьнь,евлияния
система крови (содержание эритроцитов и гемоглобина)
выделительная система
обратная афферентация
Рис. 15. Функциональная система поддержания газового
гомеостаза крови.
Особую роль в поддержании газового гомеостаза играет именно система дыхания.
Регуляция интенсивности внешнего дыхания направлена на конечный приспособительный результат - оптимизацию газового состава внутренней среды организма, т.е. чтобы легочная вентиляция
соответствовала метаболическим потребностям организма.
Основным принципом регуляции дыхания является принцип
32
саморегуляции: отклонение показателей от нормального уровня немедленно вызывает цепь процессов, направленных на их
восстановление.
Регулируемыми параметрами системы внешнего дыхания
являются глубина и частота дыхательных движений, обеспечиваемые работой дыхательных мышц, относящихся к скелетной мускулатуре.
В основном внешнее дыхание регулируется рефлекторным путем, включающим три основных звена - афферентное (рецепторы и афферентные пути), центральное (нервные центры), эфферентное (эфферентные пути и регулируемые объекты).
Центр вегетативной регуляции расположен в ретикулярной
формации продолговатого мозга. Дыхательный центр по соответствующим нервам ритмично посылает к диафрагме и межреберным мышцам импульсы, которые вызывают дыхательные движения. дыхательным центром продолговатого мозга непроизвольно регулируются частота и глубина дыхания, что обеспечивает поддержание легочной вентиляции, адекватный уровень газообмена в тканях организма. В тоже время те же дыхательные мышцы помимо сигналов от нейронов дыхательного центра получают сигналы от других нервных центров (например, рвоты, кашля и т.п.), включая корковые зоны, осуществляющие сознательный контроль деятельности
дыхательных мышц.
Кроме нервной регуляции дыхания, определенную роль выполняет
и гуморальная регуляция. Наиболее важными гормонами, участвующими в регуляции дыхания, являются адреналин, половые гормоны и др.
3.1. Характеристика дыхательного центра
Учение о дыхательном центре началось еще с Галена, наблюдавшего остановку дыхания у животного после отделения у него головного мозга от спинного. Для первого этапа развития учения особо важное значение имели наблюдения Legallois (1812) и Flourans (1842), сделавших первую попытку определить местоположение дыхательного центра. Укол иглой в область нижнего угла ромбовидной ямки продолговатого мозга мгновенно останавливал дыхательные движения
мышц туловища.
Более точную локализацию дыхательного центра изучил казанский
физиолог Н.А. Миславский (1885). Анализируя результаты перерезок, электрического раздражения различных участков продолговатого мозга, Миславский пришел к заключению, что дыхательный центр находится в ретикулярной формации продолговатого мозга по обеим сторонам шва на уровне корней подъязычного нерва и функционально состоит из
инспираторной и экспираторной части.
Поперечные перерезки ствола выше продолговатого мозга, в
частности позади бугров четверохолмия приводили к стойкому урежению дыхания (Langendorff, 1881 ), а при дополнительной двусторонней
33
ваготомии к судорожному типу дыхания. Позднее было показано, что длительные задержки на вдохе (апнейзис) не развиваются, если сохраняется передняя часть варолиева моста, так как в данной области имеется пневмотаксический центр.
По современным представлениям под дыхательным центром понимают сравнительно ограниченную совокупность нейронов в области продолговатого мозга, способных генерировать дыхательный ритм. При разрушении этой зоны ритмическое дыхание исчезает.
Электрофизиологическими исследованиями доказано, что в правой и левой половине дыхательного центра нет четкого деления на инсnираторный и экспираторный отделы. Имеются 2 скопления нейронов ретикулярной формации инсnираторных и экспираторных, импульсная активность которых меняется в соответствии с фазами дыхательного цикла (рис.16).
1
11
А 1111111111 1111111111
11111111~1\IШIII IIIIIШ~\111111
1
в
Рис. 16. Паперны импульсной активности инспираторного (А) и экспираторного (Б) нейронов в различные фазы дыхательного цикла (В).
Дыхательные нейроны располагаются в двух группах ядер - дорсальная и вентральная (рис. 17).
ДОРСАЛЬНАЯ группа ядер, располагающаяся вблизи от ядра одиночного тракта, почти на 90% состоит из инспираторных нейронов, активирующихся в фазу вдоха. Их аксоны направляются к мотонейронам ядра диафрагмального нерва в шейных сегментах спинного мозга и мотонейронам инспираторных мышц. Инспираторные нейроны получают афферентацию от механорецепторов дыхательных путей, периферических хеморецепторов и др.
ВЕНТРАЛЬНАЯ группа ядер содержит как инспираторные (в шейных сегментах С1-2), так и экспираторные нейроны (около обоюдного ядра), активирующихся в фазу выдоха. Аксоны экспираторных нейронов связаны преимущественно с мотонейронами межреберных и брюшных мышц, расположенных в грудных и поясничных сегментах спинного мозга, частично с мотонейронами диафрагмы. Инспираторные и экспираторные нейроны вентральной группы получают
34
афферентацию от инспираторных нейронов дорсальной группы, нейронов моста и др.
Кроме того, нейроны дорсальной и вентральной групп связаны реципрокными тормозными и возбуждающими связями друг с другом и некоторые из них с нейронами пневмотаксического центра.
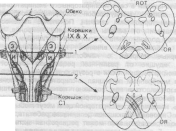
Рис. 17. Расположение инспираторных (И) и экспираторных (Э) нейронов в продолговатом мозгу кошки. Слева - дорсальная поверхность; справа - два поперечных среза, на которых изображены область скопления дыхательных нейронов (темным) и положения ядра одиночного тракта (ЯОТ) и обоюдного ядра (ОЯ). IX и Х - корешки языкоглоточного и блуждающего нервов; С1 - корешок первого шейного спинномозгового нерва.
3.2. Центральный дыхательный ритмогенез
Итак, дыхательные движения грудной клетки управляются ритмической активностью нейронов дыхательного центра. Ритмические разряды этих нейронов осуществляются автоматически, обеспечивая ритмогенез. В тоже время их активность постоянно подстраивается к изменяющимся потребностям организма под влиянием информации, поступающей от периферических рецепторов и центральных структур.
Первые опыты по изучению ритмической активности дыхательного центра были проведены И.М. Сеченовым в 1882 г. Он обнаружил ритмическую электрическую им пульсацию в изолированном продолговатом мозге лягушки. Позднее Adrian (1931) зарегистрировал ритмическую активность в изолированном продолговатом мозге золотой рыбки.
В условиях перерезки ствола мозга под варолиевым мостом и блуждающих нервов возникает длительный тетанус инспираторных мышц, что могло бы свидетельствовать о наличии непрерывной
35
активности инспираторных нейронов в норме. Однако было показано, что дыхательные нейроны не обладают пейсмекерной активностью, а ритмические залпы возникают в системе нейронов, имеющих
специфические межнейрональные связи. Инспираторные и
экспираторные нейроны имеют различную возбудимость и
рефрактерный период. В пределах каждой системы дыхательного центра, включающей в себя инспираторные и экспираторные нейроны, последние связаны между собой обширными возбуждающими и реципрокными тормозными связями. Появление активности в таких системах зависит от работы другой цепи нейронов, которые не обязательно должны быть ритмически активными, но способны оказывать облегчающее влияние на дыхательные нейроны. Инспираторные нейроны с наибольшей вероятностью получают импульсы, идущие от различных рецепторов или от нейронов ретикулярной формации. Если в результате временной суммации этих импульсов мембранный потенциал инспираторного нейрона достигнет уровня разрядки, то в нем возникнет потенциал действия, который запустит работу всей инспираторной цепи. Т.е., начать залповую активность, например, в инспираторной цепи могут любые афферентные импульсы. Самовозбуждающаяся цепь, начав работать, будет работать бесконечно. Прекратить залп может реципрокная экспираторная цепь, которая способна активироваться только после прекращения или резкого снижения активности в инспираторной цепи. Микроэлектродными исследованиями было показано (Salmoiraghi, Baumgarten, 1961 ), что во время развития залпа в инспираторном нейроне прогрессивно возрастает уровень деполяризации, порог нейрона возрастает на 2-6 мВ. Это приводит к тому, что суммация в пределах инспираторной цепи становится менее эффективной, частота импульсации снижается, и это приводит к высвобождению экспираторных нейронов, которые, активируясь, сразу же тормозят полностью инспираторные нейроны.
Таким образом, ритмическая смена вдоха и выдоха
обеспечивается циркуляцией возбуждения в дыхательных нейронах продолговатого мозга, чей объединенный импульсный паттерн вызывает вдох и выдох - колебательный дыхательный контур.
Дыхательный цикл, задаваемый центральными нервными структурами продолговатого мозга, состоит из трех фаз (D.W. Richter, 1992):
Инспираторная.
Постинспираторная (плавное снижение активности инспираторных мышц. Вследствие этого объем воздуха, поступивший при вдохе, на какое-то время задерживается, а потом пассивно выдыхается. Соответствует первой половине выдоха).
Экспираторная (соответствует второй половине выдоха).
С учетом фазности дыхательного цикла выделяют несколько типов дыхательных нейронов, для каждого из которых характерен свой рисунок
36

разрядов (табл. 4). Согласно гипотезе J.L.Feldman (1986), генератор ритма состоит из механизмов включения и последующего выключения инсnираторной и экспираторной активности:
Инсnираторная активность начинается с залпового разряда ранних инспираторных нейронов после освобождения от торможения . со стороны постинспираторных и экспираторных нейронов.
Стартовый разряд ранних инспираторных нейронов активирует полные инспираторные нейроны, которые иннервируют мотонейроны диафрагмального и межреберных нервов.
Прекращение активности ранних инспираторный нейронов приводит к активации поздних инспираторных нейронов, которые дополнительно активируют мотонейроны диафрагмального и межреберных нервов. Кроме того, они получают информацию от легочных рецепторов растяжения, которые измеряют степень растяжения дыхательных путей во время вдоха и, в связи с этим, выполняют функцию начального выключения инспирации.
Прекращение активности всех инспираторных нейронов
растормаживает постинспираторные нейроны. В момент появления залповой активности постинспираторных нейронов выключается инспираторная фаза и начинается пассивная постинспираторная фаза, т.е. первая фаза выдоха. Параллельно включаются полные экспираторные нейроны с постоянно нарастающей активностью. В это время заторможены все другие нейроны.
Прекращение активности постинспираторных нейронов приводит к активации поздних эксп_ираторных нейронов, т.е. развитию второй половины фазы выдоха и в случае форсированного дыхания активации мотонейронов экспираторных мышц.
Таблица 4
Биоэлектрическая активность
ней
Экспираторная
инспиоатооные
Полные инспирвторные
1 1 111111111111111111
Поздние инспираторные
1111111111
Постинспираторные (ранние экспираторные
1111111111111111111111111 1 1 1
Полные экспираторные
1 1 1 1111111111111111111111111111111111111111111111111111111
Поздние экспираторные
1 1 1111111111111111111
37
r-По мнению В.А. Сафонова (2000) ритмообразующей нейронной ассоциацией является 4-х нейронная система (ранние и поздние инспираторные, постинспираторные и поздние экспираторные нейроны), объединенные между собой рекурентными тормозными связями.
Важно, что возбуждающее и тормозящее взаимодействие всех типов нейронов обеспечивает ритмическую деятельность дыхательного центра.
В вентральной группе находятся также эфферентные нейроны центров блуждающего нерва, регулирующие просвет воздухоносных путей в ритме дыхательного цикла. Максимум активности этих нейронов регистрируется в конце выдоха, что ведет к сужению просвета бронхов в результате повышения тонуса гладких мышц и способствует выдоху. Минимум активности этих нейронов регистрируется в конце вдоха, что ведет к расширению просвета и облегчает вдох.
Координация деятельности правой и левой половин дыхательного центра является еще одной функцией дыхательных нейронов. Это осуществляется за счет синаптического взаимодействия различных типов дыхательных нейронов двух половин. Предположительно, этими нейронами являются экспираторные нейроны комплекса Бетцингера.
Чем круче нарастает инспираторная активность и быстрее развивается вдох, тем резче растягиваются легкие, вдох быстрее сменяется выдохом. В итоге увеличивается и глубина, и частота дыхания.
В онтогенезе спонтанная активность нейронов дыхательного центра начинает появляться в период внутриутробного развития (у человека - со 2-3-го месяцев развития), когда воздухоносные пути эмбриона еще заполнены жидкостью, а легкие находятся в спавшемся состоянии. Дыхательные движения в этом случае не выполняют дыхательной функции. Возбуждение дыхательного центра у плода является следствием пейсмекерных свойств сети дыхательных нейронов продолговатого мозга. По мере формирования синаптических связей дыхательного центра с различными отделами ЦНС пейсмекерный механизм дыхательной активности постепенно теряет свое физиологическое значение. Ритмические дыхательные движения плода становятся все более регулярными, но за сутки до родов прекращаются.
После рождения первый вдох (первый крик) происходит в момент либо пережатия пуповины, вероятно, вследствие резкой стимуляции хеморецепторов накопившимся СО2, либо вследствие раздражения проприорецепторов мышц, механорецепторов дыхательных путей и кожи, повышающих активность ЦНС, в том числе и активность центрального дыхательного механизма. Визуальный анализ родовой деятельности показал, что первое дыхательное движение ребенка - это чихание, т.е. резкое сокращение экспираторных дыхательных мышц, при
котором повышается внутригрудное давление, и производятся «взрывные» звуковые эффекты.
Чихание сопровождается выбросом слизи и прочих масс из носоглотки, носа и ротовой полости, последующей паузы, напоминающей собственно вдыхательное заполнение легких воздухом и их расправление при активном участии сурфактантов.
Вначале дыхание новорожденного носит характер судорожных вдохов, из воздухоносных путей удаляется жидкость, легкие принимают расправленное состояние, в плевральной полости формируется отрицательное давление. Затем в течение нескольких суток устанавливаются ритмичные дыхательные движения, нормализуется газовый состав и кислотно-основное состояние крови.
Итак, свойство автоматии дыхательных нейронов отличается от истинной автоматии, свойственной пейсмекерным клеткам проводящей системы сердца и . гладкой мускулатуры. Дыхательные нейроны генерируют дыхательный ритм лишь при условиях:
Сохранности синаптических связей между различными группами дыхательных нейронов;
Наличия афферентной стимуляции со стороны центральных и периферических рецепторов, среди которых особая роль принадлежит хеморецепторам;
Поступления сигналов от других отделов ЦНС, вплоть до коры.
3.3. Факторы, реrулирующие дыхание
Все афферентные факторы, влияющие на глубину и частоту дыхания, можно разделить на специфические и неспецифические (рис. 18).
Среди специфических факторов выделяют влияние:
Рсо2, Ро2, рН;
импульсации от нейронов пневмотаксического центра;
импульсации с рецепторов растяжения легких;
импульсации с проприорецепторов дыхательных мышц.
Среди неспецифических факторов выделяют влияние:
импульсации с механорецепторов легких и верхних
дыхательных путей;
импульсации с барорецепторов
сосудистых зон;
импульсации с механорецепторов кожи;
рефлексогенных
температуры тела;
гормонов и БАВ.
Особая роль отводится структурам центральной нервной системы, модулирующим активность нейронов дыхательного центра, таким как спинной мозг, мост мозга, средний мозг, ретикулярная формация, гипоталамус, кора большого мозга.
39
3.3.1. специфические факторы регуляции дыхания
Хеморецепторы
Основным регулятором активности центрального дыхательного механизма является афферентная сигнализация о газовом составе внутренней среды 'органиэма.
Впервые непосредственное влияние изменения газового состава крови на дыхательный центр продемонстрировано в опытах бельгийца Л. Фредерика (L. Fredericq) на собаках. У двух собак перерезают сонные артерии и перекрестно их соединяют. Так же поступают и с яремными венами. Позвоночные артерии перевязываются. В результате этих операций голова первой собаки получает кровь от второй собаки, а голова второй собаки - от первой. У первой собаки перекрывают трахею, что вызывает гипервентиляцию (частое и глубокое дыхание) у второй собаки, в голову которой поступает кровь от первой собаки, обедненная кислородом и обогащенная углекислым газом. У первой собаки наблюдается апноэ, в ее голову поступает кровь с более низким Рсо- и нормальным содержанием 02: гипервентиляция вымывает СО2 и практически не влияет на содержание 02 в крови, так как гемоглобин насыщен кислородом практически полностью. Результаты опыта свидетельствуют о том, что дыхательный центр возбуждается либо избытком углекислого газа, либо недостатком кислорода.
Опыты Холдена уточнили (дыхание с мешком Дугласа), что главным стимулятором дыхания является СО2, недостаток кислорода не возбуждает дыхательный центр.
Увеличение содержания СО2 в альвеолах на 0,2% ведет к увеличению вентиляции легких на 100%. При повышении Рсо2 в артериальной крови до 60 мм рт.ст. вентиляция легких возрастает до 60 л/мин. Однако при Рсо2 70 мм рт.ст. дыхание тормозится.
Важную роль в регуляции дыхания играет также рН крови. При снижении рН вентиляция легких увеличивается. В случае возрастания рН выше нормы (7,4) вентиляция уменьшается, хотя и в меньшей степени.
Сигнализация о газовом составе крови исходит от центральных (бульбарных) и периферических (артериальных) хеморецепторов.
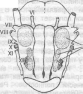
Центральные бульбарные хеморецепторы Центральные хеморецепторы расположены на вентролатеральной поверхности продолговатого мозга около корешков блуждающего и подъязычного нервов (рис. 19).
Центральные хеморецепторы в большей мере чувствительны к напряжению СО2 и рН крови и внеклеточной жидкости.
В настоящее время полагают, что основным химическим фактором, действующим на дыхание, является содержание ионов н' в межклеточной жидкости ствола мозга, а действие СО2 связано с образованием этих ионов.
41
Увеличение локальной концентрации н' оказывает влияние на дыхательный ритмогенез по сигналу от центральных хеморецепторов.
При дыхательном ацидозе повышенное Рсо, вызывает увеличение диффузии СО2 через барьер кровь - головной мозг. Повышенное СО2 приводит к росту концентрации ионов н+ около центральных хеморецепторов, которые обнаруживают это изменение и сигнализируют в дыхательный центр через повышение активности инспираторных нейронов о необходимости повышения вентиляции, компенсирующей
дыхательный ацидоз.
V
Хеморецеnторные зоны, участвующие в реrуляции дыхания
Рис. 19. Хемочувствительные зоны в продолговатом мозге кошки
Активация центральных хеморецепторов приводит к увеличению дыхательного объема и легочной вентиляции.
Угнетение центральных хеморецепторов приводит к угнетению инспираторной активности и остановке дыхания.
Периферические хеморецепторы
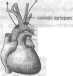
Периферические рецепторы расположены в рефлексогенных зонах - в каротидных тельцах, расположенных в области ветвления общей сонной артерии на наружную и внутреннюю сонные артерии и в аортальных тельцах (рис. 20).
~ jтельце сонной артерии
~ аортальные тельца
дуга аорты легочная артерия
Рис 20. Периферические хеморецепторы
42
Химические факторы Рсо2, Ро2 и рН влияют на минутный объем дыхания по-разному. В норме главную роль в регуляции дыхания играет
Рсо2.
Однако при утрате чувствительности центральных хеморецепторов
(хронические легочные заболевания, отравление барбитуратами) основным стимулирующим фактором спонтанного дыхания становится артериальная гипоксия, возбуждающая периферические хеморецепторы. Если такому больному дать подышать чистым кислородом, то основной стимулятор дыхания устраняется, и больной может погибнуть в результате остановки дыхания.
При нарушениях кислотно-щелочного равновесия главным
фактором становится метаболический ацидоз. Так, в ответ на снижение рН возникает гипервентиляция, выделение С02 увеличивается, и рН возвращается к нормальному уровню.
Импульсация с рецепторов растяжения легких
Роль блуждающих нервов в регуляции вдоха и выдоха доказали Геринг и Брейер в опыте с раздуванием легких воздухом в различных фазах дыхательного цикла. Оказалось, что раздувание легких воздухом тормозит вдох, после чего наступает выдох. Уменьшение объема легких (забор воздуха) тормозит выдох, ускоряет вдох. После перерезки блуждающих нервов раздувание легких не изменяет характер дыхания - тормозной эффект отсутствует, дыхание становится резко замедленным и глубоким, вдох продолжается больше обычного (как при разрушении пневмотаксического центра, наступает апнейзис).
Современные исследования выявили наличие в каждом легком более 1000 медленно адаптирующихся рецепторов растяжения, которые представляют собой чувствительные окончания толстых миелинизированных волокон блуждающего нерва и расположены в гладкомышечном слое трахеи и бронхов. Частота импульсов в рецепторах увеличивается с наполнением легких, т.е. с изменением трансмурального давления, что обеспечивает ввод информации в дыхательный центр об уровне наполнения легких.
Медленно адаптирующиеся рецепторы растяжения играют важную роль в рефлексе Геринга-Брейера, который регулирует окончание вдоха и продление выдоха. Быстрое наполнение легких во время вдоха увеличивает частоту импульсации рецепторов, что ведет к возбуждению поздних инспираторных нейронов и экспираторных, которые в свою очередь тормозят ранние инспираторные нейроны, что ведет к торможению полных инспираторных нейронов и прекращению вдоха. Т.е. прекращение вдоха наступает тем скорее, чем глубже данный вдох и чем быстрее он развивается. Рефлексы Геринга-Брейера включаются, когда дыхательные объемы превышают 1 л, при спокойном дыхании они не
проявляются.
44
Проnриоцеnтмвные афференты
Проnриорецеnторы (мышечные веретена и сухожильные
рецепторы Гольджи) в межреберных мышцах и мышцах живота могут играть существенную роль в регуляции вентиляции, особенно при ф.зоческой наrрузке. мышечные веретена иrрают важную роль в регулировании силы сокращения дыхательных мышц, особенно в -енерировании максимальной силы выдоха для устранения обструкции воздухоносных путей. Если укорочение мышцы ,_оказывается меньше заданного (например, вследствие повышенного сопротивления дыханию, имnульсация от интрафузальных волокон усиливается и через посредство у-петли повышается активность соответствующих спинальных а-мотонейронов. Сокращение мышцы усиливается, что способствует преодолению возникшего препятствия. Диафрагма - rлавная мышца вдоха, лишена мышечных волокон, но содержит
сухожильные рецепторы.
3.3.2. Несnецифические факторы регуляции дыхания
Механо е епто ы легких и ве хних ыхательных n ей Ирритаmн1:~1е рецепторы
Быстро адаптирующиеся рецепторы, также известные как uррuтатные рецепторы, составляют вторую основную категорию еrочных механорецеnторов. Чувствительные окончания толсть1х .,,.иелинизированных вагусных афферентных волокон локализованы еЖАУ эпителиальными кпетками воздухоносных путей.
Чувствительны к резкому изменению объема легких, частичкам
пыли, слизи, химическим веществам, холодному воздуху. Стимуляция быстро адаптирующихся рецепторов вызывает бронхоконстрикцию и частое поверхностное дыхание. Результатом раздражения ирритатных рецелторов является частое, поверхностное дыхание, кашлевой рефлекс, или рефлекс бронхоконстрикции. При кашле после вдоха сильно сокращаются мышцы выдоха, резко повышается внутриrрудное и внутрилеrочное давление (до 200 мм рт.ст.). открывается голосовая щель, и воздух из дыхательных путей под большим напором
высвобождается наружу и удаляет раздражающий агент.
~-
с-волокна
С-волокна - тонкие немиелинизированные волокна,
представляющие собой медленно проводящие ваrусные афференты. Окончания С-волокон расположены в паренхиме легких (легочные Сволокна), воздухоносных путей и кровеносных сосудах (бронхиальные С-волокна). Следовательно, они взаимодействуют с медиаторами и БАВ, присутствующими в крови капилляров или
бронхиальных артерий.
Активность С-волокон не зависит от фаз дыхания, им принадлежит
небольшая роль при нормальном вдохе и выдохе. Эти волокна
..,е
45
стимулируются вдыханием раздражающих агентов, а также механическим раздражением, приводя к частому поверхностному дыханию, которое ограничивает рассеивание потенциально вредных агентов по трахеобронхиальному дереву, брадикардии и секреции слизи.
В состав С-волокон входят J-рецепторы или юкстаальвеолярные рецепторы, которые расположены в паренхиме легких, прилегающей к капиллярам (юкстакапиллярам), и стимулируются развитием интерстициального отека, эмболии мелких легочных сосудов, а также при действии биологически активных веществ (никотин, простагландины, гистамин). Активация J-рецепторов вызывает закрытие гортани и апноэ, за которым следует частое поверхностное дыхание (одышка).
Рецепторы верхних воздухоносных путей
Рецепторы верхних воздухоносных путей, располагающиеся в носовой полости и верхних воздухоносных путях, отвечают как на механические, так и химические . стимулы и по механизму действия сходны с быстроадаптирующимися рецепторами в легочной паренхиме. Их стимуляция приводит к защитным рефлексам - кашлю, чиханию, бронхоконстрикции и спазму гортани. Афферентные волокна входят в состав многих нервов - блуждающего, языкоглоточного, тройничного. Чихание обусловлено раздражением рецепторов слизистой оболочки полости носа, например, пылевыми частицами или газообразными наркотическими веществами, табачным дымом, водой, в результате чего развивается глубокий сильный выдох, сужение бронхов.
Кожные и висцеральные рецепторы
Кожные рецепторы также участвуют в передаче информации в дыхательный центр, о чем свидетельствует возникающая при боли и повышении температуры кожи гипервентиляция.
Дыхательный центр реагирует и на импульсацию от сосудистых барорецепторов - повышение артериального давления приводит к торможению как инспираторных, так и экспираторных нейронов, в результате чего уменьшаются одновременно глубина и частота дыхания.
Температура тела
Повышение температуры при лихорадочном состоянии у человека сопровождается увеличением вентиляции легких. Тем самым увеличивается испарение влаги со слизистой оболочки воздухоносных путей и поверхности альвеол. Таким образом, дыхательные пути и легкие участвуют в отдаче тепла организмом во внешнюю среду, хотя тахипноэ в условиях гипертермии у человека, по сравнению с животными, не имеет существенного значения для отдачи тепла при наличии потовых желез. При этом альвеолярная вентиляция остается на уровне, соответствующем интенсивности газообмена за счет учащения дыхательных циклов, но уменьшения дыхательного объема. В результате возрастает вентиляция в основном мертвого пространства, а
46
не альвеол. Тепловая одышка опосредована локализованным в гипоталамусе центром терморегуляции, реагирующим на повышение емпературы крови, а также на информацию от периферических -ерморецепторов.
Резкое охлаждение (глубокая гипотермия) приводит к угнетению
дыхания.
Гуморальная регуляция
Кроме рефлекторных влияний на работу дыхательного центра оказывают влияние различные гормоны. Так, легочная вентиляция возрастает при поступлении в кровь адреналина (во время физической ипи умственной нагрузки), при повышении уровня прогестерона (при беременности), тиреолиберина, субстанции Р. Угнетающее влияние на активность дыхательного центра оказывает наркоз, морфин, а также эндогенные опиоиды - энкефалины и эндорфины.
3.3.3. Роль высших отделов цнс в регуляции дыхания
Центральный дыхательный механизм находится под контролем высших надмостовых (супрапонтийных) структур - мозжечка, среднего и -оомежуточного мозга, коры больших полушарий.
Известно, что человек может управлять дыханием произвольно, но до определенного уровня. Это позволяет ему менять частоту и глубину лыхания и даже задерживать его. Однако длительность произвольного апноэ ограничена: увеличение напряжения в крови СО2 при -ооизвольном апноэ приведет к сильному возбуждению хеморецепторов, ~-о выведет дыхание из-под произвольного контроля и дыхательные движения возобновятся.
Таким образом, центральный дыхательный механизм жестко граничивает вмешательство в регуляцию дыхания со стороны :;упрапонтийных структур, охраняя жизненно важную гомеостатическую, -вэообменную функцию дыхательной системы.
С другой стороны у человека не наблюдается апноэ при дыхании слородом или гипервентиляции, т.к. сказывается модулирующее злияние коры мозга.
Участие высших отделов ЦНС в регуляции дыхания человека эроявпяется и в неприятных ощущениях одышки (диспноэ ), которые возникают у него при функциональных нагрузках на кардиореспираторную систему, например, при мышечной работе,
ыхательной недостаточности.
Итак, в дыхательной системе сочетаются соматические и висцеральные структуры. Этим она принципиально отличается от других систем организма. Нормальное протекание процессов внутреннего и внешнего дыхания в различных условиях среды, их взаимодействие и координация осуществляются благодаря наличию в основном нервных регуляторных механизмов, являющихся составной частью системы
47
дыхания. Они обеспечивают передачу информации, ее переработку и посылку к эффекторам управляющих воздействий. Именно нервный аппарат соединяет отдельные участки дыхательной системы в единую функциональную систему дыхания.
3.4. Различные типы дыхания
В результате изменения дыхательного объема и частоты дыхания может значительно изменяться паттерн дыхания.
Паттерн дыхания существенно меняется при нарушении функции структур мозга, участвующих в регуляции процесса дыхания, а таюке в условиях гипоксии, гиперкапнии и при их сочетании (рис. 22).
а
б
в
Рис. 22. Патологические типы дыхания
Периодическое дыхание Чейн-Стокса (рис. 21, а). Постепенно возрастает амплитуда дыхательных движений, потом снижается и после паузы (апноэ) вновь постепенно возрастает. Возникает при нарушении работы дыхательных нейронов продолговатого мозга, а также при гипокапнии, может наблюдаться в условиях высокогорья, у здоровых людей во сне, при патологических состояниях - при отравлениях, уремии. При данном типе дыхания во время фазы глубоких дыхательных движений СО2 вымывается, и его напряжение в крови падает. В результате стимулирующее влияние СО2 на дыхательные центры практически прекращается и происходит остановка дыхания. Во время этой остановки СО2 накапливается в крови до тех пор, пока его напряжение не достигнет величины, при которой наклон кривой возрастает, вследствие этого вновь возникает гипервентиляция.
Апнейстическое дыхание (рис. 21, 6). Апнейзис - нарушение процесса смены вдоха на выдох: длительный вдох, короткий выдох и снова - длительный вдох.
Гаспинг (рис. 21, в), или терминальное редкое дыхание, которое проявляется судорожными вдохами-выдохами. Оно возникает при резкой гипоксии мозга или в период агонии. Наблюдается у недоношенных детей, при повреждении мозга.
Дыхание Биота. Проявляется в том, что между нормальными дыхательными циклам.и "вдох-выдох" возникают длительные паузы - до
48
30 с. Такое дыхание развивается при повреждении дыхательных нейронов варолиева моста, но может появиться в горных условиях во время сна в период адаптации, повышении внутричерепного давления.
Дыхание Куссмауля. Наблюдается при снижении рН крови в результате накопления нелетучих кислот (метаболический ацидоз при сахарном диабете), в результате чего возникает гипервентиляция с очень глубоким дыханием.
Апное сна. Характерны длящиеся от 1 О с до нескольких минут периоды остановки (апноэ) дыхания. В зависимости от причины выделяют обструктивное и центральное апноэ сна, которые имеют разные механизмы. Центральное апноэ сна вызвано нарушением структур ЦНС, участвующих в формировании дыхания. Обструктивное апноэ сна связано с закрытием верхних дыхательных путей.
Атактическое дыхание, т.е. неравномерное, хаотическое,
нерегулярное дыхание. Наблюдается при сохранении дыхательных нейронов продолговатого мозга, но при нарушении связи с дыхательными нейронами варолиева моста.
При дыхательной апраксии больной не способен произвольно менять ритм и глубину дыхания, но обычный паттерн дыхания у него не нарушен. Это наблюдается при поражении нейронов лобных долей озга.
При нейрогенной гипервентиляции дыхание частое и глубокое.
Возникает при стрессе, физической работе, а таюке при нарушениях структур среднего мозга.
Все виды паттернов дыхания, в том числе и патологические, возникают при изменении работы дыхательных нейронов продолговатого озга и варолиева моста. Наряду с этим могут развиваться вторичные изменения дыхания, связанные с различной патологией или воздействием на организм экстремальных факторов внешней среды. апример, застой крови в малом круге кровообращения, гипертензия алого круга или амнезия вызывают учащение дыхания (тахипноэ).
Дыхание типа Чейна-Стокса часто развивается при сердечной недостаточности. Метаболический ацидоз, как правило, вызывает брадипноэ.
3.5. Дыхание в измененных условиях
В различных условиях среды обитания системы нейрогуморальной регуляции дыхания и кровообращения функционируют в тесном взаимодействии, как единая кардиореспираторная система. Особенно четко это проявляется при интенсивной физической нагрузке и в условиях гипоксии - недостаточном снабжении организма кислородом. В процессе жизнедеятельности в организме возникают различные виды гипоксии, имеющие эндогенную и экзогенную природу.
49
Рассматривая резервные возможности дыхательной системы, следует иметь в виду, что тренировка и адаптация способны выявить, но не изменить их естественные пределы. Поэтому в экстремальных условиях существования необходимо обеспечивать поддержание таких параметров дыхательной среды, которые не выходили бы за эти границы.
3.5.1. Дыхание при физической нагрузке
Во время выполнения физической работы мышцам необходимо большое количество кислорода. Потребление 02 и продукция СО2 возрастают при физической нагрузке в среднем в 15 - 20 раз. Обеспечение организма кислородом достигается сочетанным усилением функции дыхания и кровообращения.
Было показано, что уже в начале мышечной работы, когда нет еще накопления СО2 и недоокисленных продуктов в крови, вентиляция легких быстро увеличивается. В возникновении гиперпноэ (одышки) в начале физической работы периферические и центральные хеморецепторы, как важнейшие чувствительные структуры дыхательного центра, еще не участвуют. Этот быстрый компонент дыхательной реакции на работу .объясняется главным образом нейрогенными факторами.
Прежде всего, дыхание стимулируется афферентной
импульсацией, поступающей в ЦНС из проприорецепторов работающих мышц. Кроме того, уровень вентиляции может регулироваться сигналами, поступающими к дыхательному центру главным образом из гипоталамуса, лимбической системы и двигательной зоны коры большого мозга, т.е. условно-рефлекторно. Возможно, что в стимуляции дыхания здесь участвует и выброс в кровь катехоламинов, сопутствующий повышению активности симпатической нервной системы.
По мере продолжения работы через 3-4 мин точность гипервентиляции, соответствие ее конкретным условиям адекватного снабжения газами крови, обеспечивается хеморецепторами,
контролирующими уровень PcoL-Po2. рН крови. При этом
чувствительность центральных и периферических хеморецеnторов к химическим раздражителям может изменяться. Например, под влиянием выделяющегося во время работы адреналина происходит сужение собственной артерии каротидного тельца. Снижение кровотока хеморецептора повышает его имnульсацию. Усиленная импульсация от хеморецепторов дополнительно стимулирует активность центрального механизма, в результате чего наступает компенсаторный рост вентиляции, обеспечивающей нормальный газовый состав и кислотноосновное состояние крови во время работы.
При тяжелой физической работе на уровень вентиляции оказывают влияние также повышение температуры, артериальная двигательная гипоксия и другие факторы.
so
После окончания работы легочная вентиляция быстро падает благодаря выключению нейрогенных стимулов. Однако в течение еще некоторого времени хеморецепторы стимулируются циркулирующими в крови недоокисленными продуктами обмена, в результате чего вентиляция остается повышенной. Происходит постепенное погашение образовавшегося кислородного долга, т.е. разности между общим количеством киспорода, требуемым для покрытия всех энергозатрат, и того его количества, которое было фактически потреблено за время работы.
Таким образом, наблюдаемые при физической работе изменения дыхания обеспечиваются сложным комплексом нервных и гуморальных механизмов.
3.5.2. Дыхание при гипоксии
Гипоксией (кислородной недостаточностью) называется
состояние, наступающее в организме при неадекватном снабжении тканей и органов кислородом или при нарушении утилизации в них кислорода в процессе биологического окисления (А.М. Чарной, 1961 ). В общих чертах различают:
гипоксическую гипоксию, обусловленную снижением Ро2 артериальной крови за счет уменьшения Ро2 во вдыхаемом воздухе либо при снижении давления (высотная или гипобарическая гипоксия) либо при нормобарической гипоксии;
гемическую гипоксию, обусловленную снижением кислородной емкости крови, например, при отравлении СО;
циркуляторную гипоксию, возникающую при местных
нарушениях кровообращения. В этих случаях Ро2 крови нормальное, но поступление крови к тканям нарушено;
тканевую гипоксию, возникающую при нарушении
использования тканями кислорода в дыхательной цепи при нормальном его содержании в крови.
Реакция внешнего дыхания на снижение содержания кислорода в атмосферном воздухе (гипоксическая гипоксия) зависит от продолжительности и скорости нарастания гипоксического воздействия, степени потребления кислорода (покой и физическая нагрузка), индивидуальных особенностей организма и совокупности генетически обусловленных свойств и наследственных морфофункциональных признаков.
Важнейшей срочной компенсаторной реакцией на гипоксическую гипоксию является гипервентиляция. В основе ее возникновения лежит суммарное раздражение хеморецепторов низким уровнем Ро2 (ниже 60 мм рт.ст.) и поступающими из тканей кислыми продуктами. Наблюдаемая в условиях кислородной недостаточности первоначальная гипоксическая стимуляция дыхания (гипервентиляция) приводит к вымыванию
51
углекислоты из крови и развитию дыхательного алкалоза. Гипоксия сочетается с гипокапнией. В свою очередь, это способствует уменьшению рН внеклеточной жидкости мозга и резкому снижению активности центральных хеморецепторов. Это вызывает настолько существенное торможение нейронов дыхательного центра, что он
становится нечувствительным к стимулам, исходящим от
периферических хеморецепторов. Наступает своеобразная
гипоксическая "глухота". Несмотря на сохраняющуюся гипоксию, постепенно гиперпноэ сменяется непроизвольной гиповентиляцией, что в определенной мере способствует также сохранению физиологически необходимого количества углекислоты.
Индивидуальная устойчивость человека к гипоксической гипоксии весьма вариабельна и в известной степени зависит от его тренированности. Реакция на гипоксию у коренных жителей высокогорья и у горных животных практически отсутствует, и, по мнению многих авторов, у жителей равнин гипоксическая реакция также исчезает после продолжительной (не менее 3-5 лет) их адаптации к условиям высокогорья. Однако высоту около 7-8 км, где атмосферное и альвеолярное Ро2 падают примерно втрое, считают предельно переносимой для человека.
Основными факторами долговременной адаптации к условиям гипоксической гипоксии являются; повышение содержания углекислоты и понижение содержания кислорода в крови на фоне снижения чувствительности периферических хеморецепторов к гипоксии, увеличения плотности капилляров и относительно высокого уровня утилизации тканями 02 из крови. У горцев также возрастают диффузионная способность легких и кислородная емкость крови за счет роста концентрации гемоглобина. Одним из механизмов, позволяющих горцам в условиях гипоксии повысить отдачу кислорода тканям и сохранить углекислоту, является способность повышенного образования в эритроцитах метаболита глюкозы - 2,3 дифосфоглицерата. Этот метаболит снижает сродство гемоглобина к кислороду.
Особый интерес с точки зрения теоретической и практической медицины представляет возможность использования измененной газовой среды для повышения устойчивости организма к действию неблагоприятных факторов. В условиях современной жизни человек лишен интенсивных физических нагрузок, а соответственно и естественных гипоксических стимулов. Для восполнения дефицита гипоксических регуляторов и повышения резистентности организма к стрессорным ситуациям предложено использовать барокамерные тренировки или естественный горный климат (Н.Н. Сиротинин, 1939; НА Агаджанян, 1970), вдыхание гипоксических газовых смесей при нормальном давлении (10 - 15% кислорода), либо пребывание в камере искусственного горного климата - орототерапию (В.А. Березовский, 1975; А.Я. Чижов, 1988) ..
3.5.3. Дыхание при высоком атмосферном давлении
Во время водолазных и кессонных работ человек находится под давлением выше атмосферного на 1 атм. на каждые 10 м погружения. Следовательно, на глубине 100 м человек вдыхает газовую смесь под давлением, превышающим атмосферное в 1 О раз. В этих условиях увеличивается количество газов, растворенных в крови, и особенно азота. Азот под давлением вызывает у человека наркотический эффект, кроме того, увеличивается плотность газовой смеси с азотом, что создает добавочное сопротивление дыханию.
При быстром подъеме водолаза на поверхность физически растворенные в крови и тканях газы не успевают выделиться из организма и образуют пузырьки - кровь ·"закипает". Кислород и углекислый газ быстро связываются кровью и тканями. Особую опасность представляют пузырьки азота, которые разносятся кровью и закупоривают мелкие сосуды (газовая эмболия), что сопровождается тяжелыми повреждениями ЦНС, органов зрения, слуха, сильными болями в мышцах и в области суставов, потерей сознания. Такое состояние, возникающее при быстрой декомпрессии, называется кессонной болезнью. Пострадавшего необходимо вновь поместить в среду с высоким давлением, а затем постепенно производить декомпрессию.
Вероятность возникновения кессонной болезни может быть значительно снижена при дыхании специальными газовыми смесями, например гелиево-кислородной. Гелий почти нерастворим в крови, обладает меньшей плотностью, быстрее диффундирует из тканей.
3.5.4. Дыхание при гипероксии
Высокое парциальное давление • гипероксия - может оказывать как лечебное, так и токсическое действие. Дозированное применение кислорода под давлением - гипербарическая оксигенация юпользуется для лечения некоторых состояний, связанных с тканевой и емической гипоксией. Оксигенотерапия основана на принципе
заместительной терапии: недостаток кислорода в тканях организма восполняется введением добавочного количества этого газа.
Наряду с наблюдаемым саногенным эффектом оксигенотерапии наблюдаются отрицательные реакции повреждающее действие «испорода. Дыхание чистым кислородом свыше 12-15 часов может вызвать раздражение слизистой оболочки воздухоносных путей, нарушение функции сурфактантов, а дыхание кислородом под высоким давлением (более 2-3 атм) - тяжелые расстройства функции ЦНС (судороги) уже через 1-2 часа воздействия, вазоконстрикторные реакции, нарушение микроциркуляции и аэрогематического и гематопаренхиматозного барьеров.
53
КРАТКИЙ СЛОВАРЬ БИОГРАФИЧЕСКИХ сведений и основных понятий
1771 - А. Лавуазье (А. Lavoisier, Франция) назвал газ, исчезающий при дыхании животных и человека, кислородом и показал, что углекислый газ выделяется живыми организмами в количестве,
эквивалентном потреблению кислорода.
1812 - С. Легаллуа (С. Legallois, Франция) путем перерезок доказал
местонахождение дыхательного центра в продолговатом мозгу.
1842 - Ф. Дондерс (F. Donders, Голландия) предложил
механическую модель для изучения вентиляции легких.
1859 - И.М. Сеченов (Россия) заложил основы теории газообмена в легких и транспорта газов. Сконструировал прибор абсорбциометр для
извлечения газов из крови.
1868 - И. Брейер (J. Breuer, Австрия), К. Геринг (К. Hering,
Германия) доказали, что растяжение легких является стимулом для нервных окончаний в легких, сигнал проводится по блуждающим нервам
к дыхательному центру. Это явление, основанное на
взаимосопряженных рефлексах (Геринга-Брейера), названо
«саморегуляцией дыхания».
1882 - И.М. Сеченов (Россия) впервые обнаружил ритмическую
автоматию нейронов дыхательного центра.
1882 - Н. Цунц (N. Zunz, Германия) ввел понятие «мертвое
пространство».
1885 - Н.А. Миславский (Россия) доказал, что структура
дыхательного центра продолговатого мозга состоит из инспираторного и
экспираторного отделов.
1904 - К. Бор (Ch. Bohr, Дания) впервые выявил своеобразную
форму кривых диссоциации растворов гемоглобина - их двойной изгиб. 1915- Дж. Холдейн (J. Haldane, Великобритания) определил состав альвеолярной смеси газов и вывел закономерности ее постоянства. 1920-1930-е - А. Крог, (А. Krogh, Дания) доказал, что газообмен в
легких осуществляется путем диффузии.
1926 - Э. Эдриан (E.D. Adrian, Великобритания) впервые
зарегистрировал активность нейронов дыхательного центра.
1929 - К. Нейергард (К. Neergard, Германия) установил
зависимость эластической тяги легких от поверхностного натяжения
жидкости в альвеолах.
1938 - К. Хейманс (С. Heymans, Бельгия) - Нобелевская премия за
открытие роли синусного и аортального механизмов в регуляции
дыхания.
Анатомическое мертвое пространство (МП) - объем
воздухоносных путей, в которых не происходит газообмена
Вентиляция легких - поступление воздуха в воздухоносные пути и обмен газов между альвеолами и окружающей средой
Газообмен в легких - газообмен между альвеолярным воздухом и кровью
Газообмен в тканях - газообмен между кровью и тканями организма
Диффузия - движение частиц веществ, приводящее к
выравниванию его концентрации в среде
Дыхание - совокупность процессов, обеспечивающих поступление в организм кислорода, использование его для окисления органических веществ с освобождением энергии и выделением углекислого газа в окружающую среду.
Дыхательный объем (дО) - количество воздуха, которое человек вдыхает и выдыхает при спокойном дыхании (0,5 л)
Емкость вдоха (ДО+Ровд) - максимальное количество воздуха, которое можно вдохнуть после спокойного выдоха
Жизненная емкость легких - ЖЕЛ (ДО+РОвд+РОвыд) - наибольшее количество воздуха, которое можно выдохнуть после максимального вдоха (3,0-5,0 л)
Кислородная емкость крови - количество кислорода, которое связывается кровью до полного насыщения гемоглобин
Кислородное насыщение гемоглобина (So2 или сатурация) - процент оксигемиглобина от общего содержания гемоглобина в крови
Кривая диссоциации оксигемоглобина графическая
зависимость между насыщением гемоглобина кислородом и напряжением 02 в крови
Конвекция - перенос молекул газа с потоком газовой смеси и/или жидкости
Минутный объем дыхания (МОД) - это объем воздуха, вдыхаемого или выдыхаемого за 1 мин, л/мин
Минутный объем альвеолярной вентиляции (МОАВ) - это объем воздуха, достигающего альвеол за 1 мин, л/мин
Общая емкость легких (дО+Ровд+РОвыд+ОО) - количество воздуха, содержащегося в легких на высоте максимального вдоха
Остаточный объем (00) - количество воздуха, остающееся в легких после максимального выдоха (1,0-1,5 л)
Парциальное давление газа - давление каждого газа в смеси пропорционально его доле от общего объема
Парциальное напряжение газа - давление газа в жидкости, численно равно парциальному давлению этого же газа над жидкостью в условиях равновесия
55
Паттерн дыхания - соотношение компонентов дыхательного цикла (длительность фаз, глубина дыхания)
Пневмоторакс - нарушение герметичности плевральной щели
Резервный объем вдоха (РОвд) - количество воздуха, которое человек может дополнительно вдохнуть после нормального вдоха (1,5 -1,8 л)
Резервный объем выдоха (РОвыд) - количество воздуха,
которое человек может дополнительно выдохнуть после
спокойного выдоха (1,0 - 1,4 л)
Силы поверхностного натяжения - силы поверхностного
натяжения, создаваемые водным слоем в стенках альвеол
Сурфактанты - вещества, снижающие поверхностное натяжение
Тканевое дыхание - потребление 02 тканями и выделение С02
Транспорт газов кровью - 02 от легких к тканям и СО2 от тканей
организма к легким
Транспульмональное давление
это разница между
альвеолярным и внутриплевральным давлениями
Функциональное мертвое пространство - все участки дыхательной системы, в которых не происходит газообмена
Функциональная остаточная емкость - ФОЕ (РОвыд+ОО) - количество воздуха, остающееся в легких после спокойного выдоха
Эластическая тяга легких (ЭТЛ) - упругие силы эластических элементов легочной ткани.
56
3
4
8
9
11
16
16
20
21
23
24
27
30
32
33
35
39
41
45
47
48
49
50
51
53
53
54
57
(J \
(.
ОГЛАВЛЕНИЕ
ВВЕДЕНИЕ - ..
.... ---. . (' ~ "' '
Глава 1. Эволюция дыхательнои системы :.· .
Глава 11. Этапы дыхания .
2.1. Структуры, обеспечивающие внешнее звено дыхания .
2.2. Механизм вдоха и выдоха ? ·.°'_J .
2.3. Легочная вентиляция .
2.3.1. Показатели, характеризующие легочную вентиляцию .
2.3.2. Газообмен между вдыхаемым воздухом и альвеолами .
2.4. Газообмен между легкими и кровью .
2.5. Газообмен между кровью и тканями ·.'.'·.·.· -:',,.!'.~! .
2.6. Транспорт кислорода и углекислого газа кровью _?!:·.' .. '}.:f.: ? .
2.6.1. Транспорт кислорода с -;!-.· .. J~<~ .
2.6.2. Транспорт углекислого газа r.!~~.- .. :-:".'.: .. .-: .
Глава 111. Механизмы регуляции дыхания - .
3.1. Характеристика дыхательного центра ·.;J;.,.;.i\'1;'; :: .. .'f." ..
3.2. Центральный дыхательный ритмогенез 'ч·: :.~ \~~. ::.: .. ':-: .: .
3.3. Факторы; регулирующие дыхание ~:--<·:??! .
3.3.1. Специфические факторы регуляции дыхания · .
3.3.2. Неспецифические факторы регуляции дыхания ~.:': .
3.3.3. Роль высших отделов ЦНС в регуляции дыхания :.-.-: .
3.4. Различные типы дыхания :.· .
3.5. Дыхание в измененных условиях :·:·., .':.· ;.'.-··°' .. ' .
3.5.1. Дыхание при физической нагрузi<.е :! .. :: . .' .....•. 1 ••. 1 ••..•.••
3.5.2. Дыхание при гипоксии ~.~:· .. : ..
3.5.3. Дыхание при высоком атмосферном давлении : .
3.5.4. Дыхание при гипероксии r.1.? .
КРАТКИЙ СЛОВАРЬ БИОГРАФИЧЕСКИХ СВЕДЕНИЙ ·t, ''
И ОСНОВНЫХ ПОНЯТИЙ :.~·f :~.' .. 1 ••••
ЛИТЕРАТУРА. ~ ': .. : :\.~: • • .' ~ ~ .. ~ !:? •• , •••..••...