

Билет10
1. Анаэробный путь распада углеводов (гликолиз и виды брожения). Распространение и биол. роль гликолиза. В организме человека и животных при недостаточном поступлении кислорода в клетки возможен, так называемый, анаэробный дихотомический путь катаболизма глюкозы. Так в интенсивно работающих скелетных мышцах, когда мощность механизма доставки кислорода в клетки оказываются недостаточными , включаются анаэробные механизмы синтез АТФ происходящие вне митохондрий и без участия митохондриальных ферментов. В эритроцитах , где вообще отсутствуют митохондрии , потребность в АТФ удовлетворяются только за счет анаэробного гликолиза. Интенсивный анаэробный гликолиз характерен и для опухолевых клеток.
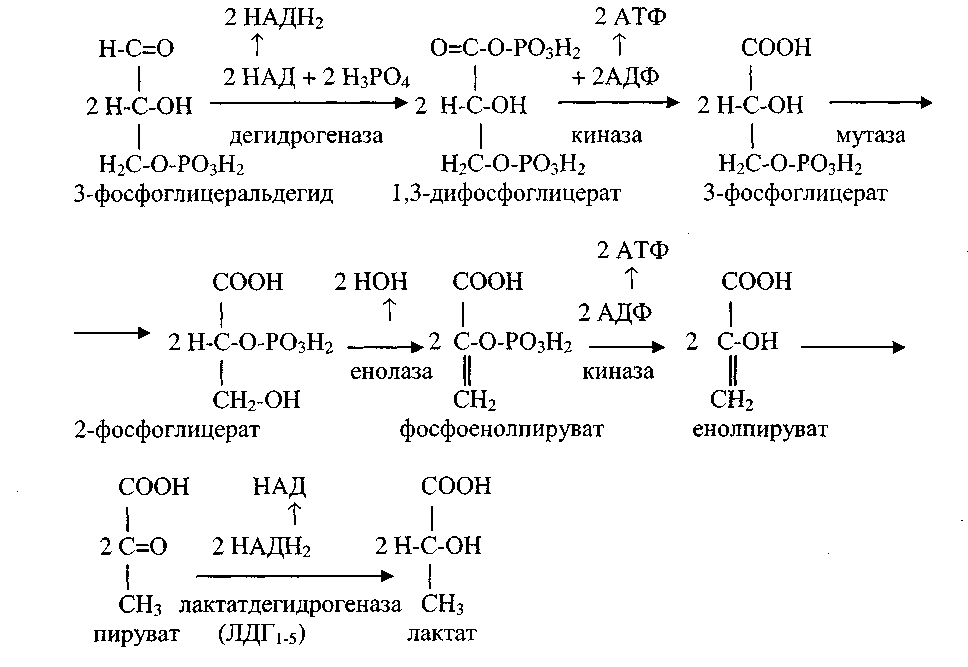
2. Челночные механизмы транспорта водорода НАДН в митохондрии.
Никотинамидные дегидрогеназы находятся не только в матриксе митохондрий, но и в цитозоле. Митохондриальная мембрана непроницаема для НАД, поэтому НАДН2, который образуется в цитозоле, может передать свой водород в митохондрию только с помощью специальных субстратных ЧЕЛНОЧНЫХ МЕХАНИЗМОВ. В митохондрию из цитозоля передается не сам НАДН2, а только водород, отнятый от него. Переносимый водород включается в молекулу вещества-челнока, способного проникать через митохондриальную мембрану. В митохондрии вещество-челнок отдает водород на митохондриальный НАД или ФАД и возвращается обратно в цитозоль.
3. Окисление субстратов и фосфорилирование АДФ в митохондриях прочно сопряжены. Скорость использования АТФ регулирует скорость потока электронов в ЦПЭ. Если АТФ не используется и его концентрация в клетках возрастает, то прекращается и поток электронов к кислороду. С другой стороны, расход АТФ и превращение его в АДФ увеличивает окисление субстратов и поглощение кислорода. Зависимость интенсивности дыхания митохондрий от концентрации АДФ называют дыхательным контролем. Некоторые химические вещества (протонофоры) могут переносить протоны и другие ионы (ионофоры) из мембранного пространста через мембрану в матрикс, минуя протонные каналы АТФ-синтазы. В результате этого исчезает электрохимический потенциал и прекращается синтез АТФ. Это называют разобщением дыхания и фосфорилирования. В результате разобщения количество АТФ снижается, а АДФ увеличивается, что приводит к возрастанию скорости окисления НАДН и ФАДН2, возрастает и количество поглощенного кислорода, но энергия выделяется в виде тепла, и коэффициент Р/О резко снижается (свободное окисление). Разобщители – липофильные вещества, легко проходящие через липидный слой мембраны: - 2,4-динитрофенол, - дикумарол (антивитамин вит. К); - билирубин (продукт распада гема); - тироксин (гормон щитовидной железы). Все эти вещества проявляют разобщающее действие только при их высокой концентрации.
4. Поэтому В3 участвует:
а) в β-окислении жирных кислот и их биосинтезе
б)в окислительном декарбоксилировании α-кетокислот (пируват, α-кетоглутарат)
в) в биосинтезе нейтрального жира, липоидов, стероидов, гема гемоглобина, ацетилхолина.
![]()
Билет13
1. Гликолиз и глюконеогенез (цикл Кори). Глюконеогенез- синтез из субстрата не углеводного происхождения.
Молочная кислота накапливается в клетках и , особенно в интенсивно работающих скелетных мышцах не яв-ся метаболитом, который подлежит выведению из организма человека. Из клеток, в которых идет анаэробный дихотомический распад глюкозы лактат поступает в кровь. Далее током крови доставляется в печень, где используется как субстрат для новообразования глюкозы – глюконеогенеза. В печени под воздействием лактатдегидрогеназы молочная кислота вначале окисляется в пируват:
Пируват в печени частично окисляется в общих путях катаболизма ,частично используется для синтеза глюкозы.
Глюконеогенез из пирувата, в принципе протекает по тому же пути, что и анаэробный гликолиз, но в обратном направлении. Однако первые две реакции глюконеогенеза отличаются от реакции гликолиза:
CH2
ǀǀ
C-O-PO3H2 фруктоза-1,6-дифосфат фруктоза-6-фосфат
ǀ фосфатаза
COOH
Фосфоенопируват
Фруктоза-6-фосфат глюкоза-6-фосфат глюкоза
Изомераза фосфатаза