

Формы бактерий с дефектом кс
|
ПРОТОПЛАСТЫ |
СФЕРОПЛАСТЫ |
L-ФОРМЫ |
|
1. Происхождение | ||
|
Из Грам+ бактерий |
Из Грам- бактерий |
Из Грам+ и Грам- бактерий |
|
2. Степень утраты КС | ||
|
Полная |
Частичная |
Полная или частичная |
|
3. Форма | ||
|
Сферическая |
Сферическая или полусферическая |
Полиморфны: крупные и мелкие сферические клетки, проходят через бактериальные фильтры. Мембранный аппарат развит хуже, чем у исходных бактерий |
|
4. Факторы, индуцирующие образование | ||
|
Обработка ферментами, разрушающими ПГ |
Обработка лизоцимом; разрушается только ПГ, НМ не разрушается |
Спонтанно, в результате действия неблагоприятных факторов (антибиотиков, лизоцима, антител) или в результате мутаций. L–трансформация может быть обратимой, если генетический контроль синтеза КС сохраняется |
|
5. Реверсия в исходную форму | ||
|
Возможна, иногда с частичной утратой физиологических признаков |
Возможна |
Нестабильные способны к реверсии в исходный вид. Устабильных неспособность к реверсии закреплена генетически |
|
6. Отношение к действию фагов | ||
|
Резистентны |
Чувствительны |
Чувствительны (нестабильные, имеющие две мембраны) или резистентны (стабильные, имеющие одну мембрану) |
|
7. Жизнеспособность | ||
|
В изотонической среде погибают в результате осмотического лизиса (плазмолиз). В гипертонической среде (когда осмотическое давление среды сбалансировано с осмотическим давлением внутри клетки) переживают некоторое время, проявляют слабую метаболическую активность, но утрачивают способность к размножению |
Способны длительно переживать в организме и на питательных средах. Могут размножаться, способы размножения более разнообразны, чем у исходных бактерий. В благоприятных условиях обладают метаболической активностью; избирательны к питательным средам, формируют на них врастающие в субстат колонии; биохимическая активность, вирулентность, антигеность ниже, а резистентность к действию антибиотиков выше по сравнению с аналогичными свойствами исходных культур | |
|
8. Выявление | ||
|
Фазовоконтрастная микроскопия |
Мало разработано, трудно поддаются лабораторной диагностике, так как морфологически неразличимы | |
|
9. Значение | ||
|
Приспособление к неблагоприятным условиям среды |
Приспособление к неблагоприятным условиям среды. В результате изменения антигенных свойств (утрата О– и К– антигенов) и снижения вирулентности способны длительно находиться (персистировать) в организме хозяина, формируя носительство бактерий (напр., при брюшном тифе) или способствуяпереходу острых заболеваний в хронические (напр., при гонорее). Становятся нечувствительными к антибиотикам, ингибирующим синтез КС | |
КИСЛОТОУСТОЙЧИВЫЕ БАКТЕРИИ
Кислотоустойчивость некоторых бактерий обусловлена особенностями химического состава их КС: наличием в ней повышенного количества липидов, оксикислот (миколовой и миколеновой), восков.
Кислотоустойчивые бактерии плохо воспринимают анилиновые красители, их окрашивают концентрированными растворами красителей с подогреванием и дифференцируют от кислоточувствительных растворами сильных неорганических кислот. Для окраски кислотоустойчивых бактерий используют метод Циля-Нильсена:
на фиксированный препарат накладывают полоску фильтровальной бумаги, наливают кислый карболовый фуксин Циля и, держа стекло над пламенем спиртовки, подогревают препарат 3–5 мин до появления паров, но не доводя до кипения;
препарату дают остыть, снимают фильтровальную бумагу, сливают краситель и промывают водой;
препарат обесцвечивают 5% раствором серной кислоты 30 сек (стекло 2–3 раза погружают в стакан с кислотой, не задерживая его в ней);
3–5 сек 5% H2SO4 или 95% C2H5OH, содержащим 3% НСl;
промывают водой;
докрашивают метиленовым синим 3-5 мин;
промывают водой, высушивают фильтровальной бумагой и исследуют с иммерсионной системой.
При обработке препарата фуксином Циля все клетки окрашиваются в рубиново-красный цвет. При последующем обесцвечивании серной кислотой кислотоустойчивые бактерии, из-за особенностей своего химического состава (высокое содержание липидов), удерживают краситель и сохраняют рубиново-красный цвет. Кислоточувствительные бактерии (напр., стафилококки и сарцины), элементы тканей и лейкоциты обесцвечиваются под действием концентрированных минеральных кислот, а при дальнейшем окрашивании метиленовым синим принимают цвет дополнительного красителя — синий (рис. 23).
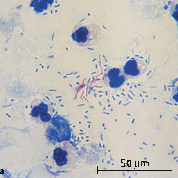
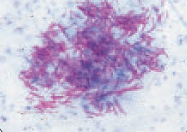
А Б
Рис. 23.Окраска по Цилю-Нильсену:
кислотоустойчивые микобактерии — красные, некислотоустойчивые бактерии, лейкоциты — синие.
А — микобактерии в моче, Б — микобактерии в мокроте
К кислотоустойчивым бактериям относятся микобактерии: M. tuberculosis, M. bovis — возбудители туберкулеза, M. africanum —возбудитель эндемического туберкулеза в Африке, M. scrofulaceum — возбудитель лимфаденитов у детей, M.leprae — возбудитель лепры, M. smegmatis — предствитель нормальной микрофлоры наружных половых органов мужчин.
ПЕРИПЛАЗМАТИЧЕСКОЕ ПРОСТРАНСТВО
Строение. Периплазматическое пространство — промежуток между ПГ КС и ЦПМ у Грам- бактерий, в котором находятся специфические белки, олигосахариды и неорганические молекулы.
Периплазматические белки представлены двумя типами: транспортными белками и гидролитическими ферментами.
Транспортные белки — это переносчики, связывающиеся с субстратами внешней среды и транспортирующие их от НМ к ЦПМ.
Гидролитические ферменты (гликозидазы, протеазы, липазы и др.) многие бактерии вырабатывают в больших количествах. Это расширяет круг используемых ими веществ. Однако гидролитические ферменты гидролизуют все типы полимерных молекул: как синтезируемые самой клеткой, так и чужеродные, попавшие в клетку извне. Отрицательные последствия гидролиза собственных молекул (самопереваривание) очевидны. Поэтому эти ферменты изолированы у Грам- бактерий от цитоплазматического содержимого в периплазматическом пространстве. Грам+ бактерии, не имеющие периплазматического пространства, выделяют гидролитические ферменты во внешнюю среду.
Функции периплазматического пространства:
Транспортная: транспорт субстратов из внешней среды к ЦПМ.
Участие в осморегуляции.
Выявление периплазматического пространства:
Электронная микроскопия (электронно-прозрачный промежуток между ПГ КС и ЦПМ).
ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНА (ЦПМ)
Строение. ЦПМ имеет толщину 5–10нм и ограничивает цитоплазму с наружной стороны. Над ЦПМ находится КС.
На долю ЦПМ приходится 8–15% сухого вещества клеток. В химическом отношении ЦПМ представляет собой липопротеин, состоящий из 15–30% липидов и 50–70% белков. Кроме того, в ЦПМ содержится 2–5% углеводов и незначительное количество РНК.
В состав мембранных липидов входят фосфолипиды и нейтральные липиды, у некоторых бактерий встречаются гликолипиды, а у микоплазм — стерины. Главным липидным компонентом бактериальных мембран являются фосфолипиды, набор которых родо- и даже видоспецифичен. Даже у одного вида бактерий липидный состав мембран непостоянен, он изменяется качественно и количественно в зависимости от условий культивирования и возраста культуры.
Все липиды бактерий — производные глицерина — содержат один или несколько остатков жирных кислот. В основном это насыщенные или мононенасыщенные жирные кислоты, полиненасыщенные жирные кислоты у бактерий отсутствуют. В составе мембранных липидов бактерий также находят кислоты, не встречающиеся в мембранах эукариот.
В структурном отношении ЦПМ — двойной слой липидов (гидрофобные концы погружены в мембрану, а заряженные гидрофильные головки торчат с двух сторон наружу) (рис. 24). Главная функция липидов — поддержание механической стабильности мембраны и придание ей гидрофобных свойств.
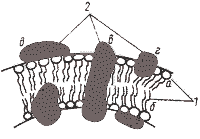
Рис. 24. Модель строения элементарной биологической мембраны:
1 — молекулы липидов:
а — гидрофильная «голова»;
б — гидрофобный «хвост»;
2 — молекулы белков:
в —интегральная;
г — периферическая;
д — поверхностная
В слой липидов включены асимметрично расположенные белковые молекулы (рис. 24). По аминокислотному составу мембранные белки не отличаются от других клеточных белков, за исключением того, что в них содержится мало цистеина. Мембранные белки разделяются на структурные и функциональные.
Структурные белки участвуют в поддержании мембранной структуры и лишены ферментативной активности. При изучении белкового состава ЦПМ не было обнаружено какого-либо универсального структурного белка. Так ЦПМ E. coli содержит 27 основных и множество минорных белков, но ни один из основных белков не присутствует в преобладающих количествах.
В отличие от липидов, у мембранных белков нет единого способа структурной организации. Для осуществления белками некоторых функций необходима их строго упорядоченная взаимная организация в мембране. Поэтому 30–50% белков имеют конфигурацию α–спирали, остальная часть белков находится в виде беспорядочного клубка.
К функциональным белкам относятся ферменты, участвующие в биосинтезе компонентов КС, окислительно-восстановительные ферменты и пермеазы, обеспечивающие перенос в клетку растворимых веществ и участвующие в процессах дыхания.
В зависимости от расположения в мембране и характера связи с липидным слоем мембранные белки делят на три группы: интегральные, периферические и поверхностные (рис. 24). Интегральные белки полностью погружены в мембрану, а иногда пронизывают ее насквозь. Связь интегральных белков с мембранными липидами очень прочна и определяется гидрофобными взаимодействиями. Периферические белки частично погружены в гидрофобную область, а поверхностные находятся вне ее. В первом случае связь с липидами в основном, а во втором — исключительно определяется электростатическими взаимодействиями. Помимо этого некоторые белки и липиды в мембране могут быть связаны ковалентно.
«Жидкая» структура мембран обеспечивает определенную свободу молекул белков, что необходимо для осуществления транспорта электронов и веществ через мембрану. Это же свойство обусловливает высокую эластичность мембран: они легко сливаются друг с другом, растягиваются и сжимаются.
В некоторых бактериальных мембранах в значительных количествах обнаружены углеводы. Они содержатся не в свободном состоянии, а входят в состав гликолипидов и гликопротеинов.
Функции мембран:
Структурная. ЦПМ — обязательный структурный элемент любой клетки. Нарушение целостности ЦПМ приводит к потере жизнеспособности клетки.
Барьерная: создает и поддерживает осмотическое давление.
Интегрирующая: ЦПМ играет координирующую роль в пространственной организации и координации ферментных систем и органелл клетки.
Транспортная. ЦПМ обеспечивает избирательное поступление в клетку веществ и ионов с использованием разных механизмов мембранного транспорта. ЦПМ является местом локализации пермеаз и окислительно-восстановительных ферментов. ЦПМ участвует в выделении из клетки продуктов обмена (токсинов и ферментов).
Энергетическая. ЦПМ участвует в процессах дыхания, так как содержит ферменты цепи переноса электронов, функционирование которых приводит к генерированию электрохимической энергии.
Участвует в биосинтезе ряда компонентов (пептидогликана КС, компонентов капсулы).
Участвует в репликации ДНК и ее последующем расхождении по дочерним клеткам.
Участвует в клеточном делении (процессе инициации и формирования поперечной перегородки).
Участвует в спорообразовании.
Выявление мембран:
1. Электронная микроскопия. ЦПМ состоит из трех слоев: двух электронно-плотных и промежуточного электронно-прозрачного.
МЕЗОСОМЫ
При избыточном росте, по сравнению с ростом КС, ЦПМ образует инвагинаты (впячивания) — мезосомы. Мезосомы — центр энергетического метаболизма прокариотической клетки. Мезосомы являются аналогами митохондрий эукариот, но устроены проще.
Хорошо развитые и сложно организованные мезосомы характерны для Грам+ бактерий. У Грам- бактерий мезосомы встречаются реже и просто организованы (в форме петли). Полиморфизм мезосом отмечается даже у одного и того же вида бактерий. У риккетсий мезосомы отсутствуют.
Мезосомы различаются по размеру, форме и локализации в клетке.
По форме различают мезосомы:
- ламеллярные (пластинчатые),
- везикулярные (имеющие форму пузырьков),
- тубулярные (трубчатые),
- смешанные.
По расположению в клетке различают мезосомы:
- образующиеся в зоне клеточного деления и формирования поперечной перегородки,
- к которым прикреплен нуклеоид;
- сформированные в результате инвагинации периферических участков ЦПМ.
Функции мезосом:
Усиливают энергетический метаболизм клеток, так как увеличивают общую «рабочую» поверхность мембран.
Участвуют в секреторных процессах (у некоторых Грам+ бактерий).
Участвуют в клеточном делении. При размножении нуклеоид движется к мезосоме, получает энергию, удваивается и делится амитозом.
Участвуют в спорообразовании.
Выявление мезосом:
Электронная микроскопия.
ЦИТОПЛАЗМА (ЦП)
Строение. Цитоплазма (протоплазма) — содержимое клетки, окруженное ЦПМ и занимающее основной объем бактериальной клетки. ЦП является внутренней средой клетки и представляет собой сложную коллоидную систему, состоящую из воды (около 75%) и различных органических соединений (белков, РНК и ДНК, липидов, углеводов, минеральных веществ).
Располагающийся под ЦПМ слой протоплазмы более плотный, чем остальная масса в центре клетки. Фракция цитоплазмы, имеющая гомогенную консистенцию и содержащая набор растворимых РНК, ферментных белков, продуктов и субстратов метаболических реакций, получила название цитозоля. Другая часть цитоплазмы представлена разнообразными структурными элементами: нуклеоидом, плазмидами, рибосомами и включениями.
Функции цитоплазмы:
1. Содержит клеточные органеллы.
Выявление цитоплазмы:
1. Электронная микроскопия.
НУКЛЕОИД
Строение. Нуклеоид — эквивалент ядра эукариот, хотя отличается от него по своей структуре и химическому составу. Нуклеоид не отделен от ЦП ядерной мембраной, не имеет ядрышек и гистонов, содержит одну хромосому, имеет гаплоидный (одиночный) набор генов, не способен к митотическому делению.
Нуклеоид расположен в центре бактериальной клетки, содержит двунитевую молекулу ДНК, небольшое количество РНК и белков. У большинства бактерий двунитевая молекула ДНК диаметром около 2 нм, длиной около 1 м с молекулярной массой 1–3х109 Да замкнута в кольцо и плотно уложена наподобие клубка. У микоплазм молекулярная масса ДНК наименьшая для клеточных организмов (0,4–0,8x109 Да).
ДНК прокариот построена так же, как и у эукариот (рис. 25).
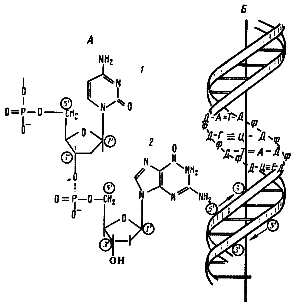
Рис. 25. Строение ДНК прокариот:
А — фрагмент нити ДНК, образованной чередующимися остатками дезоксирибозы и фосфорной кислоты. К первому углеродному атому дезоксирибозы присоединено азотистое основание:1 — цитозин;2 — гуанин.
Б — двойная спираль ДНК:Д — дезоксирибоза; Ф — фосфат; А — аденин; Т — тимин; Г — гуанин; Ц — цитозин
Молекула ДНК несет множество отрицательных зарядов, так как каждый фосфатный остаток содержит ионизированную гидроксильную группу. У эукариот отрицательные заряды нейтрализуются образованием комплекса ДНК с основными белками — гистонами. В клетках прокариот гистонов нет, поэтому нейтрализация зарядов осуществляется взаимодействием ДНК с полиаминами и ионами Mg2+.
По аналогии с хромосомами эукариот бактериальная ДНК часто обозначается как хромосома. Она представлена в клетке в единственном числе, поскольку бактерии являются гаплоидными. Однако перед делением клетки число нуклеоидов удваивается, а во время деления увеличивается до 4 и более. Поэтому термины «нуклеоид» и «хромосома» не всегда совпадают. При действии на клетки определенных факторов (температуры, pH среды, ионизирующего излучения, солей тяжелых металлов, некоторых антибиотиков и др.) происходит образование множества копий хромосомы. При устранении воздействия этих факторов, а также после перехода в стационарную фазу в клетках обнаруживается по одной копии хромосомы.
Длительное время считали, что в распределении нитей ДНК бактериальной хромосомы не прослеживается никакой закономерности. Специальные исследования показали, что хромосомы прокариот — высокоупорядоченная структура. Часть ДНК в этой структуре представлена системой из 20–100 независимо суперспирализованных петель. Суперспирализованные петли соответствуют неактивным в данное время участкам ДНК и находятся в центре нуклеоида. По периферии нуклеоида располагаются деспирализованные участки, на которых происходит синтез информационной РНК (иРНК). Поскольку у бактерий процессы транскрипции и трансляции идут одновременно, одна и та же молекула иРНК может быть одновременно связана с ДНК и рибосомами.
Кроме нуклеоида в цитоплазме бактериальной клетки могут находиться плазмиды — факторы внехромосомной наследственности в виде дополнительных автономных кольцевых молекул двунитевой ДНК с меньшей молекулярной массой. В плазмидах также закодирована наследственная информация, однако она не является жизненно необходимой для бактериальной клетки.
Функции нуклеиода:
1. Хранение и передача наследственной информации, в том числе о синтезе факторов патогенности.
Выявление нуклеоида:
Электронная микроскопия: на электронограммах ультратонких срезов нуклеоид имеет вид светлых зон меньшей оптической плотности с фибриллярными, нитевидными структурами ДНК (рис. 26). Несмотря на отсутствие ядерной мембраны, нуклеоид довольно четко отграничен от цитоплазмы.
Рис. 26.Нуклеоид стафилококка
(трансмиссивная электронная микроскопия)
Фазово-контрастная микроскопия нативных препаратов.
Световая микроскопия после окраски специфическими для ДНК методами по Фельгену, по Пашкову или по Романовскому-Гимза:
препарат фиксируют метиловым спиртом;
на фиксированный препарат наливают краситель Романовского-Гимза (смесь равных частей трех красок — азура, эозина и метиленового синего, растворенных в метаноле) на 24 часа;
краску сливают, промывают препарат дистиллированной водой, высушивают и микроскопируют: нуклеоид окрашивается в фиолетовый цвет и располагается диффузно в цитоплазме, окрашенной в бледно-розовый цвет.
РИБОСОМЫ
Строение. В отличие от клеток эукариотов рибосомы бактерий не объединены в эндоплазматическую сеть и имеют меньшую молекулярную массу.
Бактериальные рибосомы — рибонуклеопротеиновые частицы размером 20 нм, состоящие из двух субъединиц 30Sи 50S(рис. 27).
|
Малая субъединица 30S содержит 16S рРНК и 21 белок; большая субъединица 50S — 23S и 5S рРНК и 34 белка. Перед началом синтеза белка происходит объединение этих субъединиц в одну — 70S (S — константа седиментации, единица Сведберга, характеризующая скорость осаждения в центрифуге при стандартных условиях). |
|
|
Рис. 27.Бактериальная рибосома |
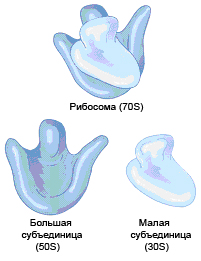
Общая масса рибосом составляет примерно 1/4 клеточной массы, а количество рибосомальной РНК (рРНК) — 80–85% всей бактериальной РНК.
Количество рибосом в бактериальной клетке зависит от интенсивности белкового синтеза и колеблется от 5000 до 90 000. Число рибосом тем больше, чем быстрее растет клетка.
В процессе белкового синтеза в клетке образуются полирибосомы (полисомы) — структуры, состоящие из большого числа рибосом, молекул информационной и транспортных РНК. Полисомы могут находиться в цитоплазме или быть связанными с мембранами клетки. По окончании синтеза белка полисомы вновь распадаются на одиночные рибосомы.
Функции рибосом:
1. Синтез белка. Существует два основных типа рибосом — 70S (у прокариот) и 80S (у эукариот). Бактериальные рибосомы, отличные от белоксинтезирующих систем эукариотических клеток, являются «мишенью» для действия многих антибиотиков. Некоторые антибиотики полностью или частично подавляют синтез белка, протекающий на рибосомах 70S, но не затрагивают функцию рибосом 80S. Так как наружная мембрана митохондрий (митохондрии содержат рибосомы 70S) очень мало проницаема, в терапевтических концентрациях антибиотики не действуют на эукариотические клетки. Деление органелл у эукариот прекращается лишь при использовании в 1000 раз более высоких концентраций антибиотиков.
Выявление рибосом:
1. Электронная микроскопия.
ВКЛЮЧЕНИЯ
Строение. У многих микроорганизмов при избытке питательных веществ в окружающей среде в ЦП клеток откладываются включения – запасные вещества разной химической природы и функционального назначения.
Включения представлены полисахаридами, липидами, полифосфатами, полипептидами, отложениями минеральных веществ (серы, кальция, железа)
(табл. 6).
Из полисахаридов в клетках откладываются гликоген, крахмал и крахмалоподобное вещество — гранулеза — специфический запасной полисахарид анаэробных споровых бактерий группы клостридий. Названные полисахариды построены из остатков глюкозы. В неблагоприятных условиях они используются в качестве источника углерода и энергии.
Липиды накапливаются в виде гранул, резко преломляющих свет и поэтому хорошо различимых в световой микроскоп. Запасным веществом такого рода является полимер β–оксимасляной кислоты, накапливающийся в клетках многих прокариот. У некоторых бактерий, окисляющих углеводороды, поли–β–оксимасляная кислота составляет до 70% сухого вещества клеток. Отложение липидов в клетке происходит в условиях, когда среда богата источником углерода и бедна азотом. Липиды служат для клетки хорошим источником углерода и энергии.
Полифосфаты, содержащиеся в волютиновых гранулах, используются клетками как источник фосфора. Полифосфаты содержат макроэргические связи и являются депо энергии, хотя их роль как источника энергии незначительна.
Все запасные вещества представлены в виде высокомолекулярных полимерных молекул, в ряде случаев отграниченных от ЦП белковой мембраной, т. е. находятся в осмотически неактивном состоянии и нерастворимы в воде.
Включения — продукты клеточного метаболизма, имеющие приспособительное значение. Когда в этих веществах возникают потребности, они снова включаются в метаболизм.
Таблица 6