

Класс II
К данному классу относятся ядерные рецепторы, не связанные с белками теплового шока. Посредством рецепторов II класса реализуют свое действие тиреоидные гормоны, ретиноевая кислота и витамин D. Рецепторы тиреоидных гормонов всегда связаны с ДНК. В отсутствие гормонов соответствующие рецепторы ингибируют экспрессию генов (рис. 20.6). Взаимодействие гормона с рецептором сопровождается образованием гетеродимерного комплекса с рецептором ретиноевой кислоты, при этом происходит снятие репрессии, присоединение коактиватора, что проявляется в индукции синтеза белка (рис.20.7).
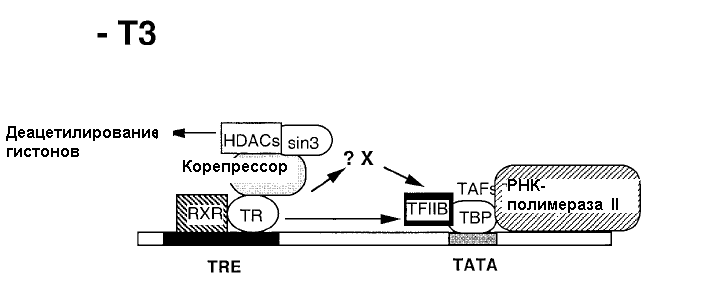
Рис. 20.6. Механизм реализация гормонального сигнала
в отсутствие йодсодержащих тиреоидных гормонов
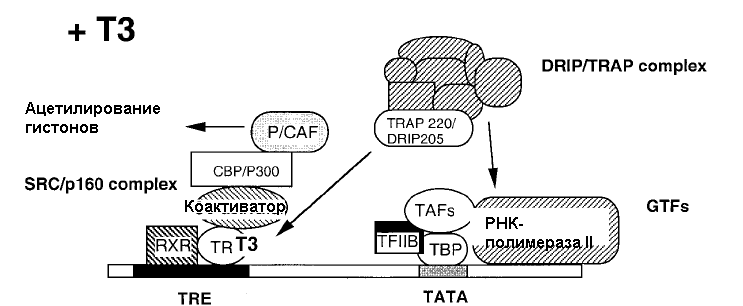
Рис. 20.7. Механизм реализация гормонального сигнала
в присутствии йодсодержащих тиреоидных гормонов
Гормоны гипоталамуса
Все гормоны гипоталамуса имеют пептидное строение и делятся на 3 подкласса:
Рилизинг-гормоны (либерины) — стимулируют секрецию гормонов передней доли гипофиза. К этой группе относятся кортиколиберин, тиреолиберин, люлиберин, фоллилиберин, соматолиберин, пролактолиберин, меланолиберин. Реализуют свой эффект посредством 7-ТМС-рецепторов, связанных с Gs-белком.
Статины — тормозят секрецию гормонов передней доли гипофиза - соматостатин, пролактостатин и меланостатин. Реализуют свой эффект посредством 7-ТМС-рецепторов, связанных с Gi-белком.
Гормоны задней доли гипофиза:
А. Вазопрессин (антидиуретический гормон) — влияющий в основном на клетки трех типов: клетки почечных канальцев, гладкомышечные клетки сосудов и клетки печени. Рецепторы АДГ в почках известны как V2-рецепторы (подробнее см. раздел «Водно-минеральный обмен»). В кровеносных сосудах и печени вазопрессин активирует V1-рецепторы, связанные с Gq-белком, активируется ФЛС, что вызывает рост внутриклеточной концентрации Ca2+ и вазоконстрикцию Таким образом, вазопрессин участвует в гомеостатическом поддержании артериального давления. В печени эффект АДГ сходен с таковым глюкагона, т.е. он стимулирует гликогенолиз и глюконеогенез.
Б. Окситоцин — действуя посредством активации 7-ТМС-рецепторов, связанных с Gq-белком, вызывает рост внутриклеточной концентрации Ca2+ и повышает сократительную активность гладкой мускулатуры матки. В лактирующей грудной железе окситоцин вызывает сокращение миоэпителиальных клеток, окружающих альвеолы и протоки молочной железы. В миндалевидном теле, вентральной части гипоталамуса, стволе мозга человека также обнаружены рецепторы к окситоцину, отвечающие за формирование полового поведения, моногамность, материнский инстинкт.
Гормоны аденогипофиза к этой группе гормонов относятся следующие гормоны белково-пептидной природы:
Кортикотропин, гонадотропины (фоллитропин и лютропин), тиреотропин реализуют свое действие через 7 тмс-рецепторы.
Пролактин и соматотропин — через 1-тмс-рецепторы, не обладающие собственной каталитической активностью.
Так, тиреотропный гормон действует на 7-ТМС-рецепторы тироцитов, активируя АЦ и ФЛС. Активация АЦ и последующее увеличение уровня цАМФ усиливают потребление йода клетками железы и повышают активность тиреопероксидазы, участвующей в биосинтезе Т3 и Т4. Активация ФЛС необходима для секреции образовавшихся гормонов в кровоток. ТТГ увеличивает также экспрессию гена гексокиназы-1. Спустя несколько часов после введения ТТГ увеличивается синтез ДНК, иРНК, ускоряется синтез белков и фосфолипидов, которые необходимы для роста и увеличения числа тироцитов и образования фолликулов. Тиреотропин воздействует и на периферические рецепторы, повышая активность селензависимой монодейодиназы периферических тканей и чувствительность рецепторов тканей к тиреоидным гормонам.
АКТГ — действует на рецепторы адренокортикальных клеток коры. Рецепторы к АКТГ — это 7-ТМС-(R), включающие механизм синтеза цАМФ. Под контролем данного механизма находится поступление в клетки холестерола в составе ЛПНП, гидролиз в коре надпочечников эфировхолестерола, транспорт холестерола в митохондрии (синтезSTAR-протеина), синтез ферментов стероидогенеза (десмолаза). Также АКТГ присуща жиромобилизующая и меланоцитстимулирующая активность.
Гонадотропины действуют на 7-ТМС-рецепторы, ассоциированные с Gs-белком, т.е. активируют АЦ систему. У женщин ФСГ стимулирует рост и созревание фолликулов яичника, а также образование рецепторов ЛГ на поверхности тека-клеток и клеток гранулезы, стимулируя таким образом синтез эстрогенов. ЛГ способствует синтезу андрогенов в тека-клетках, которые затем под действием ФСГ превращаются в эстрогены в клетках гранулезы. В больших дозах ЛГ стимулирует разрыв доминантного фолликула, т.е. непосредственно овуляцию. После овуляции ЛГ способствует развитию и поддержанию функции желтого тела и стимулирует синтез прогестерона в лютеинизированных клетках гранулезы. У мужчин ФСГ стимулирует эпителий канальцев яичка и активно влияет на сперматогенез. Он действует на клетки Сертоли и стимулирует продукцию ими белков и питательных субстратов, необходимых для созревания сперматозоидов. ЛГ инициирует развитие и созревание интерстициальных клеток (клеток Лейдига) и стимулирует биосинтез ими андрогенов.
Гормон роста (СТГ) — опосредует свое действие через 1-ТМС-рецепторы по описанному выше механизму. Срочный эффект СТГ (инсулиноподобный), реализуемый посредством IGF, проявляется в усилении липогенеза и гликолиза. Продолжительное действие СТГ, напротив, активирует липолиз и глюконеогенез, снижает поступление в клетку глюкозы, но усиливает транспорт аминокислот, активность ДНК-полимеразы, синтез белков в костях, хрящах, внутренних органах. Патология проведения гормонального сигнала: синдром Ларона – дефект рецептора к СТГ, африканские пигмеи — пострецепторное нарушение передачи сигнала.
Лактогенный гормон (пролактин) — также связывается с 1-TMS-рецептором и передает сигнал аналогично СТГ. Пролактин — это анаболический гормон, усиливающий синтез лактальбумина, казеина, триацилглицеролов, фосфолипидов и снижающий экскрецию воды. Пролактин стимулирует иммунозащитные реакции, удлиняет лютеиновую фазу менструального цикла, тормозит овуляцию и наступление беременности, путем взаимодействия с рецепторами к пролактину в клетках Лейдига регулирует в них синтез тестостерона.
ЙОДСОДЕРЖАЩИЕ ГОРМОНЫ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
В основе структуры данных гормонов лежит тирониновое ядро, состоящее из двух конденсированных молекул L-тирозина, иодированное 3 или 4 атомами йода. В фолликулах щитовидной железы образуются 2 основных гормона: Т3 (3,5,3'-трийодтиронин) и Т4 (3,5,3',5'-тетрайодтиронин).
Поступление I- в ЩЖ происходит при помощи Na+/I- симпорта, для которого используется энергия Na+/K+-АТФазы. Далее тиреопероксидаза катализирует окисление иодид-иона и присоединение иода к остаткам тирозина тиреоглобулина с образованием моно- и дийодтирозинов, которые далее конденсируются, приводя к формированию Т3 и Т4. При этом молекулы тиреоглобулина перемещаются в просвет фолликула, где накапливаются; секреция гормонов происходит при участии гидролаз лизосом эпителиальных клеток.
В периферических тканях Т4 превращается в более метаболически активный Т3 под действием селенсодержащей 5'-дейодазы. В плаценте работает 5-дейодаза и образуется малоактивный rТ3, что является механизмом защиты плода от тироксина матери.
Тиреоидные гормоны действуют на внутриклеточные рецепторы II класса по описанному выше механизму. Результатом является активация таких генов синтетазы жирной кислоты, малик-фермента, фосфоенолпируват карбоксикиназы, белков разобщения окислительного фосфорилирования, гормона роста, и в то же время, ингибирование генов пролактина, ТТГ, тиреолиберина.
Биологическим ответом организма на действие тиреоидных гормонов является усиление основного обмена (повышает потребление О2 клетками, кроме мозга, РЭС, гонад), теплопродукции на холоде, эффекта катехоламинов на сердце, моторики ЖКТ, стимуляция роста и клеточной дифференцировки. Тиреоидные гормоны необходимы для нормального развития ЦНС плода.