

Культура изолированных клеток в ткани.
Дифференциация клеток на ткани (гистогенез) и органы (морфогенез) осуществляется на основе дифференцированной работы генома, которая идет по заданной программе. В зависимости от концентрации гормональных, питательных веществ, электрических зарядов происходит дерепрессия или репрессия определенных участков генома и, как следствие, биохимическая, а затем анатомо-морфологическая дифференциация. Имеется ряд условий, способствующих этому.
1. Полярность— это свойственная растениям специфическая дифференциация процессов и структур в пространстве. При этом физиолого-биохимические или анатомо-морфологические различия изменяются в определенном направлении, в результате чего один конец отличается от другого. Явление полярности проявляется как на одной клетке, так и на ряде клеток. Так, меристематическая клетка уже поляризована благодаря своему положению: у нее есть верх и низ. И если деление пройдет перпендикулярно оси полярности, то, несмотря на одинаковое распределение наследственного материала (ДНК), дочерние клетки будут неодинаковыми по физиологическим и структурным особенностям, по факторам наследственности, расположенным в цитоплазме, и по веществам — гормонам, регулирующим активность генома. Полярно образование отдельных органов. Так, у черенков корни образуются всегда на нижнем конце. Полярность проявляется в определенной направленности роста корня и стебля, в определенном направлении передвижения веществ. Она может обуславливаться неравномерным распределением зарядов. Верхушка побега заряжена положительно по отношению к основанию, сердцевина стебля — по отношению к поверхности. Возникновение полярности может быть обусловлено разными причинами — как внешними, так и внутренними. Важно заметить, что поляризация может быть вызвана не только при неравномерном (одностороннем) воздействии того или иного фактора среды (света, температуры, земного притяжения), но и при неравномерном его восприятии. Так, под влиянием света пигменты, его воспринимающие, передвигаются к периферии цитоплазмы. Большое влияние на возникновение полярности имеет взаимодействие клеток. Возникновение полярности под влиянием окружающих клеток получило название «эффекта поля». Окружающие клетки могут оказывать эффект благодаря неравномерному химическому, механическому или электрическому воздействию.
2. Следующим фактором, имеющим значение в дифференциации клеток, являетсянеравномерное деление. При неэквивалентном цитокинезе (даже неполяризованных клеток) цитоплазматические факторы распределяются неравномерно (ядро делится как обычно), что и вызывает дифференциацию дочерних клеток. Так, при образовании устьиц делению клетки эпидермиса предшествует концентрация цитоплазмы и органелл на одной ее стороне. Затем, после обычного деления ядра, делится сама клетка. При этом образуется одна клетка меньшего, а другая большего размера. Меньшая по размеру клетка дает начало замыкающим клеткам устьиц. Клетки ризодермы также делятся неравномерно. Меньшая клетка, богатая цитоплазмой, белком, РНК, получила название трихобласта. Именно она больше не делится, а образует вырост — корневой волосок. Неэквивалентное деление наблюдается и при образовании ситовидных элементов. При этом из одной материнской клетки образуются две дочерние клетки, из которых одна дифференцируется в элемент ситовидной трубки, а другая в клетку-спутницу.
3. Существует мнение, что отдельные ткани выделяют особые морфогенетические вещества, причем источником их является, в первую очередь, меристема. Доказательством этого служат исследования Торрея, согласно которым в меристеме корня присутствует стимул, вызывающий дифференциацию проводящей системы. Согласно его данным, одним из таких веществ, вызывающих дифференциацию, является фитогормон ауксин. Взаимовлияние тканей хорошо проявляется в явлениях, получивших название гомо- и гетерогенетической индукции. При гомогенетической индукции определенная ткань вызывает образование себе подобной. Это хорошо проявляется при срастании тканей, а также при культуре изолированных тканей. В последнем случае в тканях каллуса при соприкосновении с кусочком ксилемы возникает ксилема, а при соприкосновении с флоэмой — флоэма. При гетерогенетической индукции какая-то ткань или орган блокирует образование сходной ткани или органа. Это проявляется при образовании устьиц, которые возникают только на определенных расстояниях друг от друга. Последнее важно для регуляции испарения воды и поступления С02 в листья.
4. Необходимо также отметить, что для процесса дифференциации большое значение имеют поверхностные свойства клеток, т. е. непосредственное взаимодействие — «слипание» поверхностей. Адгезия обусловлена присутствием на поверхности клеток специфических белков — лектинов, способных к обратимому связыванию с углеводами. Пектины появляются в результате избирательной экспрессии генов на разных стадиях развития клетки. Именно эти соединения обеспечивают «узнавание» и взаимодействие клеток. Так, лектин — углеводные взаимодействия лежат в основе связывания микроорганизмов-азотфиксаторов с определенным видом бобового растения. Кроме того, появились данные, что эти взаимодействия имеют значение в защите организма от болезнетворных микроорганизмов. Согласно современным представлениям, адгезия клеток играет решающую роль в морфогенезе, или образовании определенной формы того или иного органа.
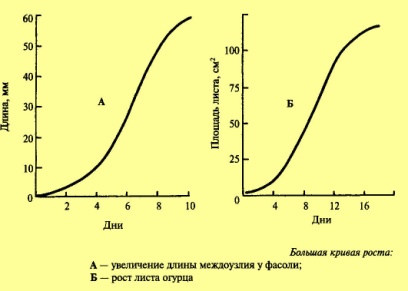
Методы измерения роста. Кривая роста по ю. Саксу. И.
Измерение
скорости роста, проведенное немецким
физиологом Ю. Саксом (1872), позволило
установить определенные закономерности.
В начальный период темпы роста, как
правило, низкие. Затем рост усиливается
и идет с большой скоростью (период
большого роста), а затем снова замедляется.
В результате рост (увеличение размера)
клетки, органа или организма в целом
может быть изображен в виде S-образной
кривой. Эта закономерность имеет
общебиологическое значение и
справедлива по отношению к росту всех
живых организмов, включая и человека.
Анализируя полученную кривую, можно ее
разделить на три участка:1) фаза, когда
рост изменяется экспоненциально
(логарифмически) ;2) фаза, когда рост
изменяется линейно;3) фаза торможения
роста. Именно период, когда рост прямо
(линейно) возрастает, называется периодом
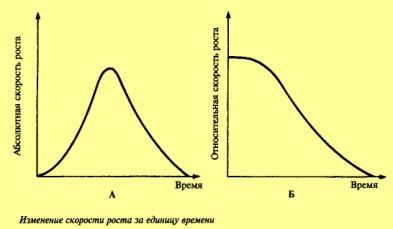
большого роста. На рисунке представлено
изменение скорости роста за единицу
времени.
 .
При этом мы видим, что вначале скорость
роста возрастает, затем некоторое время
сохраняется на постоянном уровне, а
затем падает. Падение для однолетних
растений обычно совпадает с переходом
к репродукции. Представляет интерес
изменение относительного прироста, т.
е. прироста, отнесенного к общей массе
растения. Как видим, относительные
приросты падают значительно быстрее.
Это связано с тем, что на ранних этапах
развития растение состоит почти из
одних листьев. Вырабатываемое ими сухое
вещество идет на создание все новых и
новых единиц листовой поверхности. В
результате общее количество сухого
вещества возрастает в геометрической
прогрессии. Однако затем сухое вещество
начинает все больше использоваться на
образование органов, не участвующих
в фотосинтезе. Листья составляют все
меньшую часть растения. В связи с этим,
несмотря на увеличение абсолютной
скорости прироста, его значение по
отношению к общей массе растения
постепенно падает.
.
При этом мы видим, что вначале скорость
роста возрастает, затем некоторое время
сохраняется на постоянном уровне, а
затем падает. Падение для однолетних
растений обычно совпадает с переходом
к репродукции. Представляет интерес
изменение относительного прироста, т.
е. прироста, отнесенного к общей массе
растения. Как видим, относительные
приросты падают значительно быстрее.
Это связано с тем, что на ранних этапах
развития растение состоит почти из
одних листьев. Вырабатываемое ими сухое
вещество идет на создание все новых и
новых единиц листовой поверхности. В
результате общее количество сухого
вещества возрастает в геометрической
прогрессии. Однако затем сухое вещество
начинает все больше использоваться на
образование органов, не участвующих
в фотосинтезе. Листья составляют все
меньшую часть растения. В связи с этим,
несмотря на увеличение абсолютной
скорости прироста, его значение по
отношению к общей массе растения
постепенно падает.
Влияние внешних условий на рост.
Внешние условия оказывают на рост как прямое, так и косвенное влияние. Последнее связано с тем, что скорость роста зависит от интенсивности всех остальных физиологических процессов, воздушного и корневого питания, снабжения водой, напряженности процессов обмена веществ и энергии. В этой связи влияние внешних условий может сказаться на интенсивности роста через изменение любого из указанных процессов. При этом далеко не всегда причины того или иного влияния можно с достаточной точностью установить, поскольку в естественной обстановке влияние отдельных факторов тесно взаимосвязано.
Температура.Рост растений возможен в сравнительно широких температурных границах. Растения ранневесенней флоры растут при температуре даже несколько ниже 0°С. Есть растения, для которых верхняя температурная граница роста несколько превышает 50°С. Для каждого вида растения в зависимости от его особенностей и, главным образом, от географического происхождения характерны определенные температурные границы, в которых возможно протекание ростовых процессов. Различают три кардинальные температурные точки: минимальная температура, при которой рост только начинается, оптимальная — наиболее благоприятная для ростовых процессов, и максимальная, при которой рост прекращается. Данные таблицы 7 показывают, что растения сильнее всего различаются по минимальной температуре, при которой рост начинается. Оптимальные и особенно максимальные температуры для роста различных культур очень близки. С повышением температуры от минимальной до оптимальной скорость роста резко возрастает. В области более низких температур наблюдается более быстрый подъем темпов роста при повышении температуры. Сказанное хорошо видно из данных по изменению температурного коэффициента в разных интервалах температуры. Так, скорость роста проростков гороха при повышении температуры от 0 до 10°С возрастает в 9 раз, от 10 до 20°С — в 2,5 раза, а от 20 до 30°С — всего в 1,9 раза. Оптимальные температуры могут быть неодинаковыми для роста разных органов одного и того же растения. Как правило, оптимальная температура для роста корневых систем ниже по сравнению с надземными органами. Для роста боковых побегов оптимальная температура ниже по сравнению с ростом главного стебля.Установлено, что растения интенсивнее растут в ночной период суток. Для роста многих растений благоприятной является сменная температура в течение суток — днем повышенная, а ночью пониженная. Это явление Ф. Вент назвал термопериодизмом. Явление термопериодизма хорошо проявляется на культуре томатов. Показано (Н.И. Якушкина), что пониженные ночные температуры ускоряют рост корневой системы и боковых побегов у растений. Такое влияние может быть объяснено тем, что при понижении температуры более активно работают ферменты, катализирующие распад крахмала на сахара. В листьях образуются растворимые транспортные формы углеводов, легко передвигающиеся к точкам роста корня и боковых побегов, благодаря чему их рост усиливается. Содержание воды. В процессе роста растения особенно чувствительны к недостатку воды. Уменьшение содержания воды в почве приводит, естественно, и к уменьшению содержания ее в растении, а это, в свою очередь, резко тормозит процессы роста. Снижается деление клеток и особенно их рост растяжением. Для различных физиологических процессов нужна разная насыщенность водой. Наибольшая насыщенность водой требуется для процессов роста. Насыщенность клетки или ткани растений водой называют гидратурой, она выражается в процентах. За 100%-ную гидратуру принимается такая насыщенность, при которой данное тело находится в равновесии с атмосферой, имеющей 100%-ную относительную влажность. Рост клеток идет лишь в том случае, если гидратура не падает ниже 95%. Для того чтобы поддержать такую гидратуру, точки роста надземных органов растения защищены смыкающимися листочками с хорошо развитой кутикулой. Точки роста корня не имеют подобной защиты и поэтому требуют повышенной влажности почвы для своего роста.Свет.Растения используют свет двумя путями: во-первых, как энергетический ресурс (свет служит источником энергии для синтеза органических веществ — фотосинтез); во-вторых, как сигнал или источник информации. Во втором случае энергия света может быть на несколько порядков ниже, чем в первом. Свет оказывает большое и разностороннее влияние на темпы и характер роста, как отдельных органов, так и растительного организма в целом. При этом на разные стороны ростовых процессов влияние света проявляется неоднозначно. Так, свет необходим для протекания процесса фотосинтеза, и поэтому накопление массы растения без света не идет. Вместе с тем рост клеток растяжением может идти в темноте, более того, на свету этот процесс тормозится. Свет оказывает большое влияние и на формообразовательные процессы. Этиолированные проростки, выросшие в темноте, характеризуются рядом анатомических и морфологических особенностей. В отсутствие света происходит упрощение анатомической структуры стебля. Слабо развиваются ткани центрального цилиндра, механические ткани. Вместе с тем растяжение клеток в темноте идет очень интенсивно. В результате образуются длинные, вытянутые стебли. Листья редуцированы, у двудольных растений вместо листовой пластинки образуются лишь небольшие чешуйки. Этиолированные проростки имеют слегка желтоватый оттенок. По-видимому, более быстрое вытягивание стебля и корня, этиолированных проростков выработалось в процессе эволюции, так как в большинстве случаев прорастание семян происходит в почве в отсутствие света, и эти особенности, а также отсутствие листьев облегчают проростку проникновение через слой почвы. Возможно, что вытягивание стебля в отсутствие света является следствием отсутствия ингибиторов роста. В темноте образуется много ауксинов. Нарушение соотношения ауксинов и ингибиторов вызывает несбалансированный рост. При выходе проростков на поверхность почвы происходят их внутренние и внешние изменения. В темноте у проростков двудольных растений гипокотиль изогнут, что защищает точку роста в почве от повреждений. Под влиянием света этот изгиб («крючок») выпрямляется. На свету рост стебля тормозится, рост листьев усиливается, и они принимают обычную форму. Под влиянием света происходят анатомические изменения стебля, дифференцируется эпидермис, появляются волоски, изменяется окраска — синтезируется хлорофилл. Эти изменения получили название фотоморфогенеза. Интересно заметить, что ткани проростков могут рассматриваться как «световоды», т. е. они способны проводить свет. Вследствие этого под влиянием освещения надземных органов меняются направление и темпы роста корневых систем. Именно это способствует их углублению.Изучение влияния отдельных участков спектра на перечисленные изменения (фотоморфогенез) показало, что чаще всего они вызываются при воздействии красного света с длиной волны около 660 нм. Для того чтобы свет оказал какое-либо физиологическое влияние, он должен быть поглощен каким-либо веществом. Таким веществом оказался пигмент фитохром. Было показано, что многие физиологические реакции, вызываемые облучением красным светом, можно снять при облучении дальним красным светом (длина волны около 730 нм). Эти исследования привели к заключению, что фитохром существует в двух формах, которые под влиянием облучения светом определенной длины волны могут переходить одна в другую. Фитохром, поглощающий красный свет, называют фитохром красный (Фк), а поглощающий дальний красный — фитохром дальний красный (Фдк). При поглощении красного света (660 нм) Фк переходит в Фдк, а при поглощении дальнего красного света (730 нм) Фдк переходит в Фк:может и самопроизвольно распадаться. В темноте Фдк или необратимо разрушается, или под влиянием дальнего красного света превращается в Фк. В настоящее время показано, что взаимопревращение фитохромов идет не сразу, а через ряд промежуточных форм. Надо учитывать, что на протяжении большей части дня соотношение энергии красных и дальних красных лучей составляет 3:1. Это благоприятствует превращению Фк в Фдк (активная форма). Вместе с тем в вечерние и ночные часы преобладает дальний красный свет, в связи с чем равновесие фитохромной системы сдвигается в сторону преобладания Фк (неактивная форма). Надо учитывать также, что Фк более устойчив и может синтезироваться в клетке, тогда как Фдк легко разрушается. В связи со сказанным изменения в соотношении двух форм фитохрома можно представить в виде схемы. Обе формы фитохрома выделены из растений — это хромопротеиды с молекулярной массой около 120 кДа. Хромофорная часть (собственно пигмент) представляет собой вещество, близкое по структуре к фикобилинам (красным пигментам цианобактерий и красных водорослей), состоящим из четырех пиррольньгх группировок, соединенных в открытую цепочку. Фв представляет собой восстановленную форму Фк. Фк имеет голубую окраску, а Фдк — зелено-желтую. Кроме основных линий поглощения в красной и дальней красной части спектра, оба фитохрома поглощают еще коротковолновый свет: Фк— с длиной волны 370 нм, Фдк — 400 нм. Спектрофотометрические и иммунологические исследования показали, что высокое содержание фитохрома характерно для меристематических, а также этиолированных тканей. Фитохромная система, по-видимому, очень древняя, так как она имеется даже у цианобактерий и у некоторых гетеротрофных организмов. Фитохромы можно обнаружить в различных органах растения. Физиологические проявления, которые регулируются фитохромной системой, можно отнести к фотоморфогенетическим. Основным критерием для этих реакций служит их обратимость (вызываются облучением красным светом и снимаются при облучении дальним красным светом). Особенности влияния красного и дальнего красного света видны при изучении прорастания светочувствительных семян.Рассмотрение данных в таблице показывает, что благоприятное действие на прорастание красного света снимается облучением дальним красным. К реакциям, регулируемым фитохромной системой, относятся ингибирование роста стебля, открытие крючка гипокотиля, развертывание семядолей, дифференциация эпидермиса и устьиц, образование элементов ксилемы, ориентация хлоропластов, образование антоциана, прорастание светочувствительных семян, фотопериодическая реакция растения и др. Все процессы, регулируемые фитохромной системой, делят на два типа: 1) процессы, которые под влиянием освещения красным светом усиливаются (например, дифференциация эпидермиса, синтез антоциана, прорастание семян); 2) процессы, которые тормозятся (удлинение гипокотиля, рост стебля).
Механизм действия фитохрома до настоящего времени не известен. Активной формой фитохрома является Фдк, именно его образование, которое происходит под влиянием облучения красным светом, вызывает определенный физиологический эффект. Однако и Фда не весь участвует в реакциях, а лишь его определенная часть. Возможно, что эта активная часть Фдк связана с мембранами и определенным образом ориентирована в них. В этой связи представляют интерес данные немецкого физиолога В. Гаупта, согласно которым хлоропласты ориентируются под прямым углом по отношению к лучу красного света. При этом луч может быть диаметром всего 3 мк и прямо не попадать на хлоропласты. Из этого можно сделать вывод, что фитохром локализован по преимуществу в мембранах, при этом каждая форма фитохрома определенным образом ориентирована в мембранах. При освещении определенными лучами спектра эта ориентация меняется, что и вызывает изменение ориентации хлоропластов. Воздействие Фдк может проявляться быстро (минуты) и медленно (часы). В первом случае действие Фдк, по-видимому, связано с изменением свойств мембран. Имеется предположение, что в быстрых эффектах фитохрома играют роль сигнальные цепи. Накопление Фдк в мембранах влияет на их проницаемость, в частности для К+, что, в свою очередь, изменяет электрический потенциал и вызывает определенный биологический эффект, например, никтинастическое закрывание листьев. В случае более длительных эффектов предполагают, что фитохром вызывает активацию (дерепрессию) части генома (Т. Мор). Эта точка зрения подтверждается тем, что при добавлении ингибиторов синтеза белка и РНК действие красного света не проявляется. Есть данные, что фитохром регулирует транскрипцию многих генов, связанных с позеленением, а также ген ядра, кодирующий малые субъединицы РБФ карбоксилазы/оксигеназы и связанные с хлорофиллом белки. Показано также, что красный свет индуцирует образование ряда ферментов. Ответная реакция, вызываемая Фдк или его особой формой, зависит от состояния клетки или ткани, от ее компетентности. Под влиянием красного света активность таких фитогормонов, как гиббереллины и цитокинины, возрастает. Не исключено, что действие фитохрома на геном опосредовано фитогормонами. Как уже отмечалось, многие физиологические и морфологические изменения, индуцированные фитохромом, связаны с кратковременным освещением малой интенсивности 1/100 солнечного света в течение 1 мин — низкоэнергетическими реакциями (НЭР). Однако было показано, что, для того чтобы в растениях исчезли все признаки этиоляции и они приобрели нормальный вид, этого недостаточно. Таким образом, выяснилась необходимость более длительного и более интенсивного облучения. Было сделано заключение, что такие эффекты включают высокоэнергетические реакции (БЭР). Именно БЭР обеспечивают нормальный рост побегов. При этом оказалось, что спектр действия БЭР также несколько отличен. Наибольшее влияние оказывают не красные, а дальние красные (710—730 нм) и синие лучи. Что касается пигментов (фоторецепторов), которые ответственны за эти реакции, то для проявления действия дальнего красного света это тот же Ф. Фоторецептор для синего света окончательно не установлен, возможно, что это флавиновые соединения, например, флавинокаротиноиды. В последнее время действию синего света на растения уделяется большое внимание. Показано, что синий свет влияет на электрические и генетические процессы, изменяет метаболизм. Причем его действие отличается от эффектов красного света. Примерами процессов, регулируемых синим светом, являются фототропизм, биосинтез пигментов и др. Установлена стимуляция синим светом разрушения крахмала и биосинтеза малата в замыкающих клетках устьиц. Обогащение осмотически действующими веществами приводит к открытию устьиц. Синий свет влияет на устьичные движения также путем активации Н+-АТФазы плазмалеммы. Синий свет вызывает возникновение разности потенциалов, что влияет на поступление ионов. В случае стимулирования генов синий свет стимулирует транскрипцию и трансляцию и приводит к морфогенетическим эффектам. Рассматривая воздействие света, необходимо остановиться на влиянии круглосуточного освещения на процессы роста. Опыты по выращиванию растений при свете электрических ламп (светокультура) показали, что рост многих растений при круглосуточном освещении идет значительно интенсивнее, особенно при правильном подборе качества света, т. е. типа ламп (Б.С. Мошков). Так, при выращивании сеянцев древесных культур (дуб, сосна) на непрерывном освещении темпы их роста возрастают в 1,5—2 раза (В.М. Леман). Такие однолетние растения, как горох и фасоль, также характеризуются очень интенсивным ростом в условиях круглосуточного освещения. Однако существуют растения, на рост которых круглосуточное освещение оказывает отрицательное влияние. В некоторых случаях круглосуточное освещение вызывает явления, сходные с теми, которые обычно являются следствием недостатка света. Такие растения, как томаты, в условиях непрерывного освещения вытягиваются, листья становятся желтыми, хлорофилл разрушается. Это явление называют зеленой этиоляцией. Особенно вредное влияние круглосуточного освещения проявляется при высокой ночной температуре.
Снабжение кислородом.Процессы роста требуют затрат энергии, источником которой служит процесс дыхания. В этой связи понятна необходимость кислорода. При снижении концентрации кислорода ниже 5% рост тормозится. Это происходит не только из-за нарушения энергетического баланса, но и в силу накопления продуктов анаэробного обмена (спирт, молочная кислота). Минеральное питание. Для нормального протекания ростовых процессов необходимо достаточное снабжение всеми необходимыми минеральными элементами. Особенно специфична роль снабжения растений азотом. Это связано не только с тем, что азот входит в состав белков и нуклеиновых кислот, но и с образованием двух основных групп гормонов, регулирующих ростовые процессы (ауксинов и цитокининов).