

Тема 4. Ядро клетки. Деление клетки
![]()
4.1. Клеточное ядро
![]()
4.1.1. Общие представления о составе и функциях ядра
![]()
Ядро – важнейшая структура клетки, содержащая наследственный материал – молекулы ДНК.
![]()
4.1.1.1. Ядерная днк и хромосомы: введение
I. Количество тех и других
|
ДНК |
В ядре любой диплоидной по ДНК соматической клетки человека присутствует
|
|
Хромо- сомы |
а) Каждая из молекул ДНК связана с определёнными белками, образуя дезоксинуклеопротеидный тяж – хромосому. б) Таким образом, хромосом в ядре диплоидной клетки человека – тоже 46. |
|
Наборы ДНК и хромосом |
Наборы хромосом (и содержащихся в них молекул ДНК) во всех диплоидных клетках организма
|
II. Интерфазные хромосомы
|
Опреде- ление |
а) В неделящейся или только готовящейся к делению клетке хромосомы морфологически неразличимы, т.к. находятся большей своей частью (или целиком) в деконденсированном состоянии. б) Такие хромосомы называются интерфазными. |
|
Сохранение основных свойств |
а) Тем не менее, и в этом состоянии они сохраняют
б) Так, каждая хромосома крепится к внутренней ядерной мембране обоими своими концами и рядом промежуточных участков. Причём,
|
III. Метафазные и анафазные хромосомы
|
Визуали- зация |
а) В процессе деления клетки (митоза или мейоза) хромосомы резко конденсируются, в результате чего значительно
б) После этого они становятся видимыми даже на светооптическом уровне. |
|
Различие формы |
При конденсации разные хромосомы приобретают разную форму и по этому признаку могут быть идентифицированы. |
|
Опреде- ление |
а) Морфологию хромосом, как правило, описывают по их состоянию на стадии метафазы или анафазы митоза. б) Соответственно, хромосомы на этих стадиях называются метафазными и анафазными. |
|
Гомология хромосом |
а) Изучение этих хромосом, в частности, показывает, что
б) Очевидно, это относится и к интерфазным хромосомам. |
|
Подробней структуру хромосом мы рассмотрим позже.
4.1.1.2. Характеристика хромосомной ДНК I. Строение |
|
Двойная спираль |
Любая из молекул хромосомной (ядерной) ДНК – очень протяжённая линейная структура, состоящая из двух полинуклеотидных цепей, которые
|
|
Компле- ментар- ность |
а) Комплементарность означает, что
б) В этих пaрах (А-Т и Г-Ц) нуклеотиды как бы дополняют друг друга по своей пространственной конфигурации, что обеспечивает максимально плотную упаковку цепей в двойной спирали. |
|
Длина |
а) Молекулы хромосомных ДНК существенно различаются
б) Поэтому различаются по длине и хромосомы – как интерфазные, так и метафазные. в) Средняя же длина одной молекулы ДНК человека –
г) Следовательно, общая протяжённость всех 46 молекул ДНК, находящихся в хромосомах одной клетки, – примерно 185 см. |
|
Объясне- ние высокой длины ДНК |
Такая огромная (для молекулярного уровня) длина обусловлена тем, что совокупность внутриядерных молекул ДНК (а значит, и ядро каждой клетки) содержит информацию обо всём организме человека, а именно о
всех его систем и органов. |
|
II. Информационная функция ядерной ДНК |
|
Информа- ция в ДНК |
Конкретно, в наборе молекул ядерной ДНК каждой клетки содержится следующая информация:
|
|
Этапы передачи инфор- мации |
Передача информации о структуре белка включает 3 этапа:
|
|
Строение РНК |
Напомним: РНК любого вида (мРНК, рРНК, тРНК) представляет собой одиночную цепь, состояшую из рибонуклеотидов – адениловых, уридиловых, гуаниловых и цитидиловых. |
|
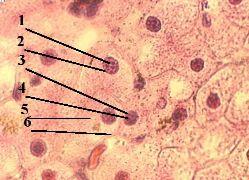
III. Обнаружение ДНК в ядрах клеток |
|
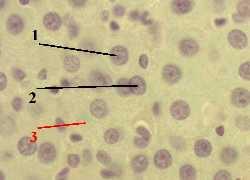
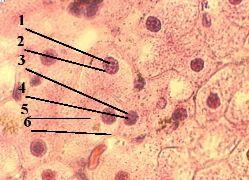
1. а) Обнаружить ДНК в клеточных ядрах можно с помощью метода Фёльгена (п. 1.1.4). – б) При этой окраске
2. а) На снимке мы видим, что, действительно, в ядрах (1) клеток содержится ДНК. б) Исключения составляют ядрышки (2):
|
1. Препарат - дезоксирибо- нуклеиновая кислота (ДНК) в ядре клетки. Окраска по методу Фёльгена.
Полный размер |
|
4.1.1.3. Функции ядра и внутриядерные процессы |
|
Из вышесказанного ясно, что ядро соматической клетки должно выполнять 2 ключевые функции:
|
|
I. Использование информации ДНК |
|
Основные процессы |
В рамках реализации данной функции в ядрах происходят указанные ранее процессы: 1. транскрипция определённых участков ДНК (синтез пре-мРНК), 2. созревание (процессинг) мРНК, 3. синтез и созревание тРНК и рРНК. |
|
Суть процес- синга |
а) Пре-мРНК обычно содержат
б) Поэтому одно из событий созревания пре-мРНК – это т.н. сплайсинг:
в) Затем к концам этой цепи присоединяются дополнительные последовательности, которые необходимы
|
|
Другие процессы в ядре (той же направ- ленности) |
К прочтению информации ДНК имеют отношение также следующие внутриядерные процессы: 4. формирование субъединиц рибосом (из рРНК и поступающих из цитоплазмы рибосомных белков) и 5. разнообразные регуляторные процессы (в том числе модификация ДНК и хромосомных белков), изменяющие активность разных участков ДНК. |
|
II. Сохранение и воспроизведение наследственного материала Эту функцию ядра также обеспечивает целая серия процессов. |
|
1. Репарация ДНК |
а) Репарация ДНК – это обнаружение и исправление возникающих повреждений структуры ДНК. б) Репарация совершается практически постоянно, но её эффективность
|
|
2. Реплика- ция ДНК |
а) Репликация (удвоение) ДНК происходит перед любым делением клетки (кроме второго деления мейоза). б) В продуктах репликации – дочерних молекулах ДНК
Это означает, что репликация является |
|
3. Конден- сация хромосом |
Конденсация хромосом
|
|
III. Уничтожение ядром своего наследственного материала |
|
а) И ещё очень важный процесс может быть инициирован в ядре:
б) Это имеет место в ходе апоптоза – клеточного “самоубийства”, которое запускается
в) Более подробно апоптоз рассматривается в конце этой темы |
|
IV. Особенности процессов в ядрах созревающих половых клеток В ядрах созревающих половых клеток тоже могут происходить все перечисленные процессы – но с рядом особенностей. |
|
1-2. Репарация и апоптоз |
а) В указанных клетках значительно ужесточаются (по сравнению с соматическими клетками) требования к качеству наследственного материала, передаваемого потомству. б) Поэтому, как считают, резко повышаются
|
|
3. Конден- сация хромосом |
а) При созревании сперматозоидов происходит гораздо более значительное уплотнение хромосом, чем при делениях клеток. б) Такая сверхконденсация хромосом достигается |
|
4-5. Конъюга- ция и кросс- синговер |
а) Кроме того, добавляются новые процессы – конъюгация и кроссинговер. б) Они совершаются в ходе особого деления – мейоза – и заключаются в том, что гомологичные хромосомы
в) Данный обмен повышает разнообразие генетического материала в половых клетках. |
|
Таким образом, процессы, происходящие в клеточных ядрах, очень разнообразны и исключительно важны.
4.1.1.4. Выявление транскрипции в клеточных ядрах Все вышеназванные процессы могут быть выявлены с помощью специальных методик. Продемонстрируем это на примере транскрипции ДНК, т.е. матричного синтеза РНК. I. Принцип метода |
|
Мечение уридином |
а) Экспериментальным животным in vivo вводят в кровь раствор радиоактивного уридина. б) Данное соединение в клетках превращается в Н3–УТФ (уридинтрифосфат) - один из четырёх нуклеотидов, используемых при синтезе РНК. в) Поэтому вскоре после введения метки она оказывается в составе новосинтезированных цепей РНК. Замечание. - При синтезе ДНК вместо уридилового нуклеотида используется тимидиловый; так что Н3–УТФ включается только в РНК. |
|
Последу- ющие проце- дуры |
а) Через определённое время животных забивают и готовят срезы изучаемых тканей. б) Срезы покрывают фотоэмульсией. В местах нахождения радиоактивного соединения
Т.е. последние являются маркёрами радиоактивной метки. в) Затем срез (после промывки и закрепления) красят как обычный гистологический препарат. |
|
II. Препарат |
|
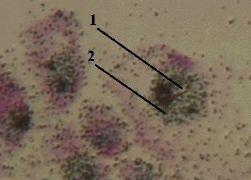
2. Препарат - включение Н3–уридина в РНК. Окраска гематоксилин-эозином. |
|
|
1. а) На представленном снимке мы видим, что меченое вещество (2) сосредоточено, главным образом, в ядрах (1) клеток. б) Это и отражает тот факт, что
|
Полный размер |
|
2. Наличие метки в других частях препарата объясняется, например, тем, что
|
|
|
4.1.1.5. Структурные компоненты интерфазного ядра |
|
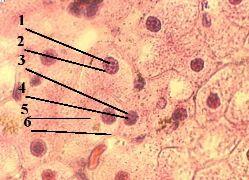
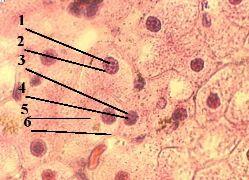
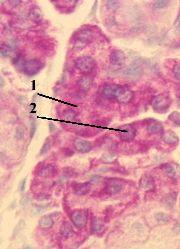
3. Препарат - структура клеточного ядра. Клетки печени. Окраска гематоксилин-эозином. |
|
|
Теперь обратимся к строению интерфазного ядра. а) Будем использовать обычный препарат печени. б) В печёночных клетках хорошо видны округлые ядра (1), окрашенные гематоксилином в фиолетовый цвет. в) В ядрах же выявляется ряд структур. |
Полный размер |
|
1. Хроматин |
а) В соответствии с вышеизложенным,
|
|
б) Поэтому используется термин "хроматин": им обозначают
Морфологически хроматин проявляется в виде тёмных глыбок (3). |
|
|
в) Однако в этих глыбках – не весь хроматин:
|
|
|
2-3. Прочие различи- мые элементы |
Кроме глыбок хроматина, в ядре можно видеть |
|
|
4-5. Матрикс |
а) Хроматин и ядрышки находятся во внешне бесструктурной среде – ядерном матриксе. б) Оказалось, что в последнем имеются
в) Иногда под термином “кариоплазма” понимают весь ядерный матрикс. |
|
|
Теперь рассмотрим перечисленные компоненты ядра более подробно.
4.1.2. Хроматин (хромосомы) Хроматин занимает основную часть объёма ядра.
4.1.2.1. Типы хроматина I. Эу- и гетерохроматин |
|
Электронная микрофотография - ядро плазматической клетки. |
|
|
1. Электронный уровень На электронных микрофотографиях обнаруживается, что хроматин подразделяется на два типа:
|
Полный размер |
|
|
|

|
2. Природа эу- и гетеро- хроматина |
а) Из предыдущего изложения ясно, что
б) В связи с этим,
|
|
3. Световой уровень |
а) При световой микроскопии эухроматин, как уже отмечалось, в ядре не выявляется. б) Те же глыбки, которые обычно обозначаются как “глыбки хроматина”, представляют собой на самом деле
|
|
4. Актив- ность эу- и гетеро- хроматина |
а) В конденсированных участках хромосом ДНК недоступна для разнообразных внутриядерных ферментов – в том числе для ферментного комплекса, осуществляющего транскрипцию ДНК. б) В связи с этим,
|
|
II. Виды гетерохроматина |
|
Опреде- ления |
а) При изменении функционального состояния клетки или в процессе её дифференцировки возможен переход части гетерохроматина в эухроматин и обратно. б) В связи с этим, гетерохроматин подразделяется на два вида:
|
|
Природа конститу- тивного гетеро- хроматина |
а) Вторая фракция гетерохроматина - это, видимо, те области хромосом, в которых ДНК
б)Данные области расположены, в основном, вблизи центромерных участков хромосом. |
|
III. Состояние хроматина в разных клетках |
|
а) Из предыдущего следует: чем больше в ядре доля гетерохроматина,
б) Для иллюстрации этого положения рассмотрим два снимка. |
|
|
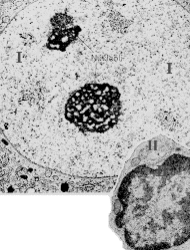
Электронные микрофотографии - I - ядро нервной клетки, II - ядро малого лимфоцита. |
|
|
1. Сверху – ядро нервной клетки (I). Здесь гетерохроматина очень мало - следовательно, ядро и клетка в целом функционально очень активны. 2. Внизу – лимфоцит (II). а) В его ядре преобладает гетерохроматин. б) Это вполне коррелирует с очень малым объёмом цитоплазмы, которая к тому же бедна органеллами. в) Данная клетка циркулирует в крови, и процессы синтеза РНК и белков идут в ней с небольшой скоростью. |
Полный размер |
|
IV. Половой хроматин (тельце Барра) Одним из компонентов гетерохроматина может быть т.н. половой хроматин (тельце Барра), встречающийся только у женщин. |
|
Половые хромосомы у мужчин |
а) У мужчин в наборе хромосом каждой клетки содержится, как известно, по одной Х- и Y-половой хромосоме. б) Обе они находятся в деконденсированном состоянии, т.е. входят во фракцию эухроматина. |
|
|
Половые хромосомы у женщин |
а) У женщин в клетках содержатся по две Х-хромосомы. б) А. Одна из них деконденсирована. Б. Вторая же Х-хромосома всегда находится в конденсированном состоянии, образуя в ядре
|
|
|
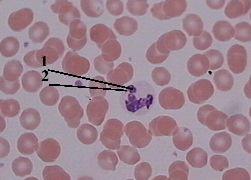
4. Препарат - половой хроматин в лейкоцитах (мазок крови женщины). Окраска азур 2-эозином. |
||
|
Обнаруже- ние полового хроматина в мазке крови |
а) Для обнаружения полового хроматина обычно исследуют мазок крови. б) В нейтрофильных лейкоцитах женщин половой хроматин выявляется |
Полный размер |
|
в) По этому признаку в судебной медицине отличают кровь женщин от крови мужчин. |
||
|
Другой способ |
Другой часто используемый в тех же целях объект –
|
|
|
4.1.2.2. Химические компоненты хроматина. Белки хромосом I. Общая характеристика |
|
Перечень компо- нентов |
Как уже отмечалось, каждая из 46 хромосом представляет собой дезоксирибонуклеопротеин (ДНП), т.е. комплекс
|
|
РНК хромосом |
а) Что касается РНК, то это
б) В последнее время особенно подчёркивается регуляторное влияние (на активность генов) т.н. малых молекул РНК. |
|
Белки |
Содержание же белков в хромосомах (и в хроматине в целом) намного выше содержания не только РНК, но и ДНК:
|
|
Охарактеризуем хромосомные белки подробней. По своим физико-химическим свойствам они делятся на основные и кислые. II. Основные (в физико-химическом смысле) белки хромосом |
|
Виды гистонов |
а) Основные белки представлены гистонами пяти видов: б) Всего на них приходится 60-80% массы всех хромосомных белков. |
|
Особен- ности состава |
Гистоны обогащены
|
|
Свойства |
При этом
|
|
Функция |
а) Указанные взаимодействия приводят к образованию
б) Таким образом, гистоны выполняют важную структурообразующую функцию. |
|
III. Кислые белки хромосом |
|
Кислые белки чрезвычайно разнообразны: имеется, по крайней мере, |
|
|
Структур- ные белки |
По всей видимости, некоторые кислые белки тоже играют структурную роль, участвуя в образовании
|
|
Ферменты |
а) Другую группу составляют многочисленные ферменты, обеспечивающие процессы
б) Это не означает, что все работающие на ДНК ферменты постоянно связаны с ней: какие-то ферменты могут находиться в кариоплазме. |
|
Регуля- торные белки |
а) Наконец, самой разнообразной по составу, видимо, является группа регуляторных белков. б) Эти белки контролируют
|
|
4.1.2.3. Деконденсированные интерфазные хромосомы (эухроматин): нуклеосомный уровень организации Как уже было сказано, благодаря гистонам хромосомы имеют нуклеосомную организацию. I. Нуклеосома |
|
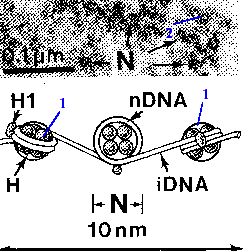
Электронная микрофотография и схема - нуклеосомы. |
|
|
а) Основа нуклеосомы –
содержащая по 2 молекулы гистонов четырёх видов (Н2А, Н2В, Н3 и Н4). б) Вокруг одной такой глобулы
что и образует в итоге нуклеосому. |
Полный размер |
|
в) В участках между глобулами с ДНК связано ещё
г) С учётом этих соединительных (линкерных) отделов, период нуклеосомной организации составляет примерно
|
|
|
II. Хромосома в целом |
|
Кол-во нуклеосом |
Молекула ДНК участвует в образовании очень большого числа нуклеосом (в среднем, 600.000). |
|
|
ДНК - цепь "бусин" |
В результате, на данном уровне организации
|
|
|
Влияние нуклеосом на доступ- ность ДНК |
а) Нуклеосомная организация не препятствует действию на ДНК внутриядерных ферментов. б) Хотя не исключено, что в момент прохождения ферментного комплекса по какому-либо участку ДНК последний
Но затем эта связь быстро восстанавливается. |
|
|
Новообра- зуемые цепи ДНК |
Новообразуемые участки строящейся цепи ДНК (при репликации ДНК) тоже
|
|
|
Итоги |
а) Таким образом, нуклеосомный уровень укладки присущ хромосомам
б) Заметим также: по сравнению с молекулой ДНК, нуклеосомная нить оказывается
благодаря закручиванию ДНК вокруг каждого октамера. |
|
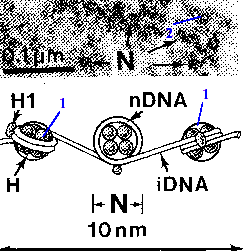
|
4.1.2.4. Конденсированные интерфазные хромосомы (гетерохроматин): нуклеомерная и хромомерная организация а) Те интерфазные хромосомы или их части, которые образуют гетерохроматин, тоже имеют нуклеосомную организацию. б) Однако здесь добавляются и следующие уровни укладки хромосомы. I. Второй уровень: нуклеомерный |
|
Роль гистона Н1 |
Данный уровень образуется (при определённых условиях) в результате взаимодействия друг с другом
|
|
Названия структуры |
При этом нуклеосомная нить конденсируется в более плотную структуру толщиной около 30 нм:
|
|
Описание структуры |
а) Полагают, что нуклеомерная нить представляет собой суперспираль, причём такую, в которой вновь (как в нуклеосомной цепи) чередуются
б) Иначе говоря,
|
|
Блокиро- вание активности генов |
а) Однако в этой цепочке
б) Поэтому данный уровень организации отсутствует у эухроматина. |
|
Сокраще- ние длины хромосомы |
Там же, где нуклеомерная структура имеется, длина хромосомы (или её отдела, если конденсируется только он)
|
|
II. Третий уровень: хромомерный |
|
Факторы и принцип укладки |
а) Дальнейшая компактизация хромосом происходит под влиянием уже не гистонов, а
б) И начинает использоваться другой принцип укладки:
|
|
Описание структуры |
а) Так,
б) Каждую розетку составляет несколько соседних петель, которые прикрепляются своими основаниями к общему белковому центру. в) Подобные центры в большом количестве содержатся в ядерном матриксе. |
|
Названия элементов структуры |
Таким образом, на этом уровне организации хромосома представляет собой цепь, состоящую из
|
|
Особен- ность гетеро- хроматина |
В гетерохроматине розетки, видимо, расположены более-менее рыхло, т.е.
|
|
4.1.2.5. Метафазные хромосомы: высшие степени конденсации I. Третий уровень: хромонемный |
|
Особен- ность метафаз- ных хромосом |
При формировании метафазных хромосом третий уровень компактизации выглядит несколько иначе:
|
|
Названия структуры |
В таком состоянии хромомеры формируют нить толщиной 300 нм:
|
|
II. Четвёртый уровень: хроматидный |
|
Вероятный способ организа- ции данного уровня |
а) Хромонема спирализуется или тоже складывается в петли. б) При этом петли, возможно, образуют группы, которые
в) Это соответствует тому обстоятельству, что под действием повреждающих факторов хромосома может распадаться на сегменты. |
|
Название структуры |
Продукт данного (и последнего) этапа конденсации –
|
|
Параметры хромосомы |
а)Толщина всей хромосомы в состоянии максимальной компактизации – вдвое больше: 1400 нм б) Зато длина хромосомы человека за счёт всех уровней укладки сокращается, в итоге, в 10.000 раз:
|
|
III. Резюме: уровни укладки метафазных хромосом Вышеперечисленные уровни укладки метафазных хромосом суммированы в следующей таблице. |
|
Уровень укладки |
Пояснение |
Толщина |
Сокращение длины |
|
I. Цепь нуклеосом |
В каждой нуклеосоме молекула ДНК делает по 2 оборота вокруг белкового октамера. |
10 нм |
В 6,2 раза |
|
II. Нуклеомерная нить (хроматиновая фибрилла, или нить) |
Это цепочка нуклеомеров, содержащих по 5-8 нуклеосом, закрученных в суперспираль. |
30 нм |
В 6-7 раз (всего - примерно в 40 раз) |
|
III. Хромонема (хромосомная фибрилла) |
Хроматиновая нить образует петли, собирающиеся в розетки, плотно прилегающие друг к другу. |
300 нм |
Примерно в 250 раз (всего - в 10000 раз) |
|
IV. Хроматида |
Хромонема складывается в петли протяжённостью в 1 сегмент хроматиды. |
700 нм |
|
|
Метафазная хромосома |
Хромосома состоит из двух связанных друг с другом хроматид. |
1400 нм |
Результи- рующее сокращение длины - как в хроматидах. |
|
4.1.2.6. Кариотип человека I. Введение |
|
Опреде- ление |
а) Завершая конденсацию к началу метафазы митоза, хромосомы принимают определённую форму. б) Соответственно, вводят понятие “кариотип” – это
|
|
Отличие анафазных хромосом от мета- фазных |
а) Заметим: метафазные хромосомы состоят из двух хроматид, которые начали расходиться, но ещё связаны в центромерной области. Поэтому они
б) Анафазные же хромосомы – это уже разошедшиеся хроматиды и потому похожи на
|
|
Набор анафазных хромосом здорового человека а) Для характеристики кариотипа используют наборы как метафазных, так и анафазных хромосом. б) В частности, на приведённом рисунке изображены анафазные хромосомы человека. |
|
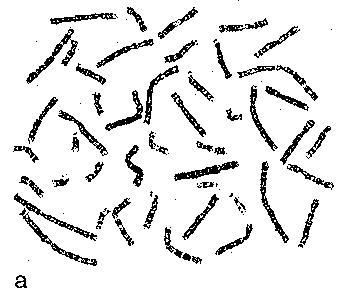
|
II. Общая характеристика кариотипа |
|
Набор анафазных хромосом здорового человека а) На этом рисунке те же хромосомы человека представлены уже в сгруппированном виде. б) Это значительно упрощает характеристику кариотипа. |
|
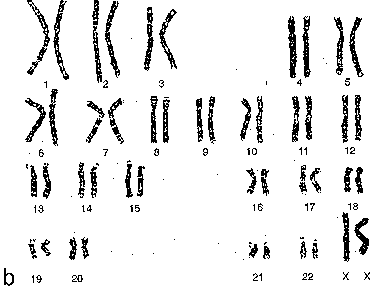
|
Пары хромосом |
а) Всего, как отмечалось в п. 4.1.1.1,III, в соматической клетке человека – 46 хромосом. б) Они попарно гомологичны друг другу, так что всего получается 23 пары. |
|
Аутосомы и половые хромосомы |
а) Хромосомы 22-х пар называются аутосомами, а одну пару образуют половые хромосомы (п. 4.1.2.1.IV):
б) При этом Y-хромосома почти вдвое короче Х-хромосомы. |
|
Группы хромосом |
Кроме того, все хромосомы по размеру и форме подразделяются на 7 групп. |
|
III. Характеристика хромосом |
|
Отделы хромосомы |
а) Почти у каждой хромосомы обнаруживаются следующие части:
б) В области центромеры находится
|
|
Морфоло- гические типы хромосом |
По положению центромеры хромосомы делят на 3 вида:
|
|
Локализа- ция генов рРНК |
а) У некоторых хромосом в одном из плеч имеется и вторичная перетяжка. б) Это место расположения генов рибосомных РНК. |
|
Набор анафазных хромосом человека с синдромом Дауна а) При т.н. хромосомных болезнях кариотип отличается от нормального. б) Например, при болезни Дауна в ядрах всех клеток присутствует дополнительная 21-я хромосома. |
|
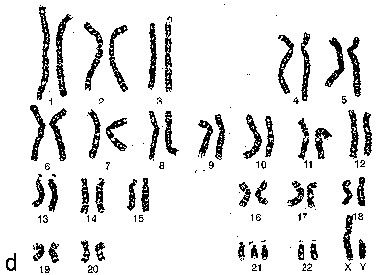
|
4.1.3. Прочие структуры ядра
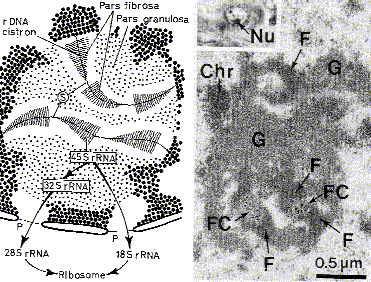
4.1.3.1. Ядрышко I. Введение |
|
Общий вид |
а) Ядрышко (или нуклеола) – самая плотная структура ядра. б) Обычно оно имеет округлую форму. в) Нередко в ядре содержится несколько ядрышек. |
|
Компоненты ядрышка При электронной микроскопии в области ядрышка выявляются 3 компонента: возле ядрышка - а в самом ядрышке -
Рассмотрим их подробней. |
Электронная микрофотография - ядрышко.
|
|
II. Ядрышковые организаторы |
|
Опреде- ление |
Ядрышковые организаторы - это те участки хромосом, которые содержат
|
|
Гены рРНК |
а) Напомним: всего имеется 4 вида рРНК (п. 3.3.1.1). б) Гены трёх из них (28S, 18S и 5,8S) расположены рядом друг с другом, образуя кластер, транскрибируемый как единое целое. в) Ген четвёртой, самой короткой, РНК (5S) располагается и транскрибируется отдельно. |
|
Количе- ство и локализа- ция генов рРНК |
а) Указанный кластер, а также ген 5S-рРНК,
б) Таким образом, всего имеется 10 ядрышковых организаторов. в) Вместе они содержат у человека,
|
|
Количе- ство ядрышек |
а) Количество ядрышек в ядре зависит от того, во сколько групп объединяются 10 ядрышковых организаторов – в одну, в две или больше. б) Таким образом, ядрышко – это не самостоятельное образование, а производное определённых хромосом. |
|
III. Фибриллярный и глобулярный компоненты |
|
Фибрил- лярный компонент |
а) На кластерах генов рРНК активно происходит синтез предшественника рРНК – единой пре-рРНК, включающей нуклеотидные последовательности трёх рРНК. б) Цепи предшественника тут же (в ядрышке) подвергаются созреванию:
в) Одновременно образуется и созревает пре-5S-рРНК. г) Эти полинуклеотидные цепи: пре-рРНК и зрелые рРНК - и составляют фибриллярный компонент ядрышек. |
|
Глобу- лярный компонент |
а) Тут же, в ядрышке, зрелые рРНК связываются с рибосомными белками, формируя субъединицы рибосом. б) Последние представляют собой глобулярный компонент ядрышек. в) Сформированные субъединицы выходят затем из ядра в цитоплазму. |
|
IV. Выявление ядрышек при световой микроскопии |
|
Окраска гем.-эоз. |
В случае световой микроскопии ядрышки (4) обычно различимы даже при обычной окраске (гематоксилином и эозином). |
|
|
5. Препарат - РНК в цитоплазме и ядрышках клеток (поджелудочная железа). Окраска по Браше (метиловым зелёным - пиронином). |
|
|
Окраска по Браше а) Можно использовать также гистохимическую реакцию на РНК (по Браше). б) Как мы видели в предыдущей теме, из всех компонентов ядра заметную реакцию при данной методике дают только ядрышки (2):
|
Полный размер |
|
в) Очевидно, это связано с высоким содержанием в ядрышках рРНК и их предшественников. г) Напомним: этот же препарат иллюстрирует и наличие РНК в цитоплазме (1) (в составе рибосом), о чём говорилось в теме 3. |
|
|
4.1.3.2. Ядерная оболочка |
|
Ядерная оболочка имеет 2 особенности:
|
|
I. Мембраны ядерной оболочки |
|
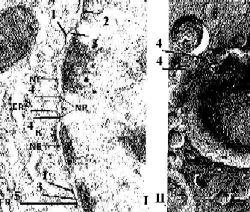
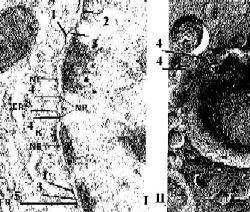
Электронные микрофотографии - ядерная оболочка. I. Обычный способ приготовления препарата. II. Метод замораживания и скалывания. |
|
|
Общее описание а) Различают
б) Они разделены перинуклеарным пространством (3). в) Таким образом, ядерная оболочка – это полый двуслойный мешок. |
Полный размер |
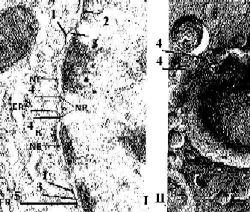
|
Природа внешней мембраны |
а) С внешней ядерной мембраной со стороны гиалоплазмы связаны рибосомы (5). б) Т.е. эту мембрану можно рассматривать как
|
|
Роль внутренней мембраны |
а) А внутренняя ядерная мембрана связана с элементом ядерного матрикса – ядерной пластинкой (ламиной). б) К последней же крепятся концы всех хромосом, причём, в строго определённых местах. в) Таким образом, внутренняя мембрана выполняет опорную функцию. |
|
II. Ядерные поры |
|
Функция |
Через поры в ядерной оболочке происходит интенсивный транспорт молекул и крупных частиц: а) в ядро из цитоплазмы – б) из ядра в цитоплазму – многочисленных
|
|
|
Коли- чество пор |
а) Поэтому количество пор в ядерной оболочке тем больше, чем интенсивней идут в клетке синтетические процессы. б) Обычно всего в оболочке ядра – 2000 – 4000 пор. |
|
|
Отверстие поры |
а) В области краёв поры (4) внутренняя и наружная мембраны сливаются, образуя округлое
б) В отверстие встроен т.н. комплекс поры. |
|
|
Комплекс поры |
а) Комплекс поры включает:
б) Из этих гранул
в) Центральная гранула связана фибриллами с периферическими. г) В итоге структура напоминает велосипедное колесо. |
|
|
Переме- щение веществ через пору |
а) Относительно небольшие молекулы диффундируют пассивно через гидрофильные каналы поры. б) А более крупные молекулы и частицы переносятся путём активного транспорта с затратой энергии ГТФ. Причём, в этом участвуют
в) По-видимому, центральная гранула – это и есть проходящий через пору транспортный комплекс. |
|
|
Специаль- ный метод исследо- вания |
а) Чтобы наблюдать внутреннюю поверхность ядерных мембран (II), используют особый способ приготовления препарата:
|
|
|
б) В итоге, ядерные поры проявляются как округлые углубления. |
||
|
4.1.3.3. Ядерный матрикс (кариоскелет) |
|
Компо- ненты |
К кариоскелету относятся
|
|
Ядерная ламина |
а) Ядерная ламина образована многочисленными промежуточными филаментами. б) Как уже было сказано, она
|
|
Фибрил- лярная сеть |
Полагают, что внутриядерная фибриллярная сеть - тоже (как и ламина) своего рода каркас, который используется для фиксации
|