

1) Аглютиногенів в оболонці еритроцитів;
- аглютиногени – речовини мукополісахаридної природи;
- мають антигенні властивості, тобто при попаданні в організм, який не має цього аглютиногена, зумовлює утворення імунних антитіл взаємодія їх з антигенами (аглютиногенами) еритроцитів злипання (аглютинація) гемоліз;
- присутні у всіх групових системах крові.
2) Аглютинінів (аглютиніни українською – злипні!) в плазмі крові;
- аглютиніни – антитіла до відповідних аглютиногенів еритроцитів;
- нормальні (природні, вроджені) аглютиніни є в системі АВ0. В більшості інших групових систем крові нормальних антитіл (аглютинінів) немає, але при попаданні в організм людей, які не містять певного аглютиногена цього аглютиногена, організм відповідає виробництвом імунних антитіл.
Групову належність необхідно враховувати при переливанні крові. Кров донора (людина, у якої беруть кров для переливання) та реципієнта (людина, якій переливають кров) мають бути сумісними. Це означає, що плазма крові реципієнта не повинна містити аглютинінів до аглютиногенів еритроцита донора.
В системі АВ0 виділяють 4 групи крові (за наявністю аглютиногенів А, В, 0 в оболонці еритроцитів та аглютинінів α та β в плазмі крові):
|
Група крові |
Наявність аглютиногенів |
Наявність аглютинінів |
|
I |
0 |
α та β |
|
II |
А |
β |
|
III |
В |
α |
|
IV |
А, В |
відсутні |
З таблиці видно, що якщо в еритроцитах є певний аглютиноген, то в плазмі крові відсутній однойменний аглютинін. Якщо в еритроцитах немає аглютиногена, то вплазмі крові є відповідний аглютинін. Це пов’язано із здатністю однойменних аглютинінів та аглютиногенів (А та α, В і β) взаємодіяти між собою. При такій взаємодії відбувається злипання еритроцитів утворення конгломератів еритроцитів закупорка судин цими конгломератами порушення мікроциркуляції. При таких реакціях виділяються біологічно активні речовини, які пошкоджуть структуру та функцію судин. Найбільше страждають судини нирок та функції органа. Аглютиновані червоні кров’яні тільця потім руйнуються (гемолізуються).
Визначення групової належності крові (групи крові) в системі АВ0 базується на використанні цоліклонів анти-А та анти-В. Цоліклони містять чисті антитіла до аглютиногенів А (цоліклон анти-А) чи В (цоліклон анти-В). На чистій сухій поверхні змішують цоліклони з кров’ю (у співвідношені 10:1) та дивляться чи зумовлює цоліклон аглютинацію еритроцитів. Якщо певний цоліклон зумовлює реакцію аглютинації, то в еритроцитах крові є відповідний аглютиноген, якщо ж не зумовлює, то це означає, що відповідного аглютиногена в еритроцитах немає. Так взнають, які аглютиногени є в еритроцитах, а яких немає та роблять висновок про групову належність крові.
|
Група крові |
Цоліклон анти-А |
Цоліклон анти-В |
|
0 (I) |
“-” немає А |
“-” немає В |
|
А (II) |
“+” є А |
“-” немає В |
|
В (III) |
“-”немає А |
“+” є В |
|
АВ (IV) |
“+” є А |
“+” є В |
“-” – немає аглютинації еритроцитів;
“+” – є аглютинація еритроцитів.
Визначення групововї належності крові необхідне перед переливанням крові. Прокуратурою дозволяється переливання лише одногрупної крові за системою АВ0!!! При переливанні неодногрупної крові може відбуватися зустріч однойменних аглютиногенів та аглютинінів реакція аглютинації еритроцитів закупорка судин та виділення біологічно активних речовин порушення функції нирок та інших органів. Такий стан має назву гемотрансфузійного шоку. В результаті такого шоку люди нерідко гинуть.
Антигени А неоднорідні. Кров більшості людей другої та четвертої груп містять сильний антиген А. В крові приблизно 12 % людей другої та четвертої груп наявний менш сильний антиген А1. Є також інші різновидності антигенів А: А2, А3, А4, А5, А6...Ах. Пам’ятати про це необхідно тому, що при наявності в еритроцитах різновидностей аглютиногена А, в плазмі крові можуть бути так звані екстрааглютиніни (α1, α2 і т. д.). При цьому люди можуть мати однакову групу крові за системою АВ0, але їх кров буде несумісна при переливанні.
Наприклад:
друга група за системою АВ0, А (ІІ) β може мати такі варіанти:
- підгрупа А1 (ІІ) β
- підгрупа А2 (ІІ) β, α1.
Кров обох людей має другу групу крові за системою АВ0, але при переливанні призведе до ускладнень (відбудеться контакт однойменних аглютиногенів та аглютинінів аглютинація еритроцитів).
Примітка: аглютиноген 0 деколи називають аглютиногеном Н – це речовина, з якою розвивається у одних людей аглютиноген А (друга група), а у інших – аглютиноген В (третя група), у третіх – А та В (четверта група). У людей першої групи аглютиногени А та В не розвиваються, в оболонках еритроцитів міститься аглютиноген 0 (Н).
Проби, що проводять перед переливанням крові.
1. Визначення групової належності донора та реципієнта за системою АВ0 (принцип – дивися вище). Дозволяється переливання тільки одногрупної крові за системою АВ0.
2. Визначення резус-належності крові донора та реципієнта (принцип – дивися нижче). Дозволяється переливання лише одногрупної крові за системою резус.
Окрім основних групових систем крові (АВ0 та резус) є ще біля 20 неосновних (М, S, КК...). За основними системами обов’язково визначають належність крові. Несумісність крові донора та реципієнта за неосновними груповими системами виключають за допомогою проб на сумісність: плазма крові реципієнта не повина містити аглютиніни до аглютиногенів еритроцитів крові донора (тому змішують плазму реципієнта з кров’ю донора у співвідношені 10 : 1).
3. Проба на індивідуальну групову належність крові донора та реципієнта. Проводять шляхом змішування плазми реципієнта з кров’ю донора на чистій, сухій поверхні при кімнатній температурі без додавання колоїдів. При таких умовах реагують повні антитіла (холодова аглютинація). Облік реакції проводять за відсутністю чи наявністю аглютинації. Її наявність свідчить про несумісність крові донора та реципієнта – переливання проводити неможна.
4. Проба на резус-сумісність крові донора та реципієнта. Проводять шляхом змішування плазми реципієнта з кров’ю донора, додають колоїд (желатин, альбумін) та ставлят на водяну баню (48-49 градусів). При таких умовах реагують неповні антитіла; часто такими антитілами є антитіла до резус-фактора (звідси назва проби). Облік реакції проводять за відсутністю чи наявністю реакції аглютинації. Її наявність свідчить про несумісність крові донора та реципієнта – переливання крові проводити неможна.
5. Біологічна проба – реципієнту тричі вводять кров внтрішньовенно струйно по 5 мл з інтервалами 5-10 хв. В інтервалах між вливанням порцій крові цікавляться станом реципієнта (головна біль, дрижаки, біль у кістках, особливо в попереку – нирки!). Якщо ці скарги не з’являються, переливають всю дозу крові.
75.Физиологическая характеристика Rh-системы крови. Значение системы Rh-при беременности и переливании крови.
Резус система, як і система АВ0, є основною груповою системою крові. Резус система влаштована відносно простіше, ніж система АВ0. Вона має лише аглютиніни, які містяться в оболонках еритроцитів. Позначаються ці аглютиногени як Д, С, Е (класифікація Фішера) чи Rh°, rh', rh'' (класифікація Вінера). Найбільш важливим (сильним) та поширеним аглютиногеном системи резус є Д (Rh°). Якщо в оболонках еритроцитів є ці аглютиногени, вона називається резус-позитивною, якщо їх немає, кров називається резус-негативною. 85% людей є резус-позитивними, 15% – резус-негативними.
Для донорів критерії більш суворі: кров донора рахується резус-негативною, якщо немає жодного аглютиногена системи резус (Rh°, rh', rh'').
В системі резус є ще три аглютиногена, які мають невелику антигенну силу та вивчені менше. Це аглютиногени с, d, e (класифікація Фішера) чи Hr°, hr', hr'' (класифікація Вінера).
Таким чином, в резус-системі відсутні природні аглютиніни, але при попаданні в організм резус-негативних людей резус-позитивної крові (може бути при вагітності резус-негативної жінки резус-позитивним плодом; при переливанні резус-негативним людям резус-позитивної крові) відбувається вироблення імунних антитіл до резус-фактора. Ці антитіла зумовлюють аглютинацію еритроцитів (плід – при вагітності, донора – при переливанні крові).
Перше переливання резус-позитивної крові резус-негативному рецепієнту не зумовлює розвиток гемотрансфузійного шоку: антитіла утворюються повільно, їхня концентрація (титр) в плазмі крові для аглютинації стане достатньою лише через 2 тижні, коли еритроцити донора вже будуть зруйновані. Але друге переливання резус-позитивної крові резус-негативній людині миттєво зумовить розвиток шоку (організм реципієнта сенсибілізований – містить достатню кількість антитіл в плазмі крові до резус-фактора імунна реакція гемотрансфузійний шок).
Тому при переливанні крові обов’язково визначають резус-належність крові донора до рецепієнта та переливають лише однгрупну кров.
Принцип визначення резус-належності крові такий самий, як і групової належності за системою АВ0. Але використовують імунні анти-резус сироватки (містять імунні антитіла до резус-фактора). Якщо сироватка під час змішування з дослідною кров’ю зумовлює реакцію аглютинації еритроцитів, значить вони (еритроцити) містять резус-фактор (кров резус-позитивна). Якщо сироватка не зумовлює реакцію аглютинації. Отже, в оболонці еритроцитів немає резус-фактора й кров резус-негативна. Дозволяється переливання лише одногрупної крові за системою резус.
При вагітності небезпека резус-конфлікта є, якщо резус-негативна породілля та резус-позитивний плід. В цих умовах організм матері виробляє імунні антитіла до резус-фактора еритроцитів плода. Як правило, перша вагітність закінчується успішно. Причина цього: плацента при нормальному протіканні вагітності не пропускає еритоцити. Тому імунізація матері під час вагітності неможлива. Плацентарний бар’єр порушується еритроцити плода попадають в організм матері імунізація матері вироблення антитіл до резус-фактора, які будуть перебувати в крові до кінця життя матері.
Під час другої вагітності антитіла до резус-фактора будуть з організму матері попадати в організм плода (ці антитіла відносяться до імуноглобулінів класу G та легко проходять через плаценту). Попавши в організм плоду, антитіла, зумовлюють аглютинацію та гемоліз його еритроцитів. При других пологах резус-позитивна кров плода знову попадає в організм матері імунізація підсилюється погіршується прогноз наступної вагітності. Таким чином, кожна наступна вагітність резус-негативної жінки резус-позитивним плодом погіршує прогноз наступної вагітності.
76. ПД типичных кардиомиоцитов желудочков, механизмы происхождения, физиологическая роль. Изобразите схему соотношения во времени ПД и одиночного сокращения міокарда.
Природа автоматії полягає в наявності в А-КМЦ специфічних потенціал-чутливих каналів. Ці канали змінюють свій стан, коли в ході реполяризації мембрани КМЦ (кінець попереднього ПД) мембранний потенціал досягає 60 mV. При цьому в мембрані А-КМЦ:
- відкриваються повільні кальцієві канали – йони кальцію за градієнтом концентрації починають повільно входити в клітини;
- відкривається повільні натрієві канали – йони натрію за градієнтом концентрації починають повільно входити в клітини;
- закриваються калієві канали – зменшується вихід калію з клітини за градієнтом концентрації.
Така зміна стану каналів мембран А-КМЦ веде до повільного зменшення мембранного потенціалу (деполяризація мембрани). Ця деполяризація виникає без дії зовнішнього подразника (автоматично), і коли вона досягає критичного рівня (– 45 mV), виникає пік ПД. Ця частина змін в часі мембранного потенціалу клітини, що володіє автоматією, є специфічною для неї і носить назву фази повільної діастолічної деполяризації, або спонтанної деполяризації.
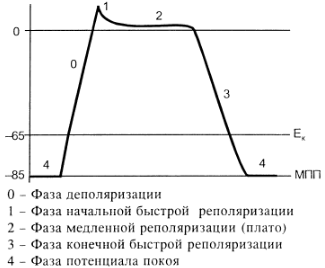
Частота, з якою центр автоматії генерує ПД, залежить від двох факторів:
1) величина порогового потенціалу; чим вона більша, тим частота менша; в звичайних умовах під впливом механізмів регуляції частіше змінюється рівень мембранного потенціалу спокою зміна порогового потенціалу зміна частоти генерації імпульсів збудження водієм ритму зміна частоти серцевих скорочень;
2) швидкість повільної діастолічної деполяризації (ПДД); механізми регуляції змінюють проникність відповідних каналів зміна швидкості ПДД зміна частоти генерації ПД водієм ритма серця зміна ЧСС.
Центри автоматії другого та наступних порядків генерують імпульси збудження рідше, ніж водій ритма (пазухово-передсердний вузол), перш за все тому, що у них менша швидкість ПДД.
Дуже рідко (в умовах патології) здатність до автоматії проявляється в Т-КМЦ (з яких побудований робочий міокард шлуночків та передсердь). Це відбувається при різкому зменшенні рівня мембранного потенціалу спокою цих клітин (до –60 mV, як в А-КМЦ; в звичайних умовах рівень їх потенціалу спокою дорівнює –90 mV). Такі центри автоматії носять назву ектопічних.
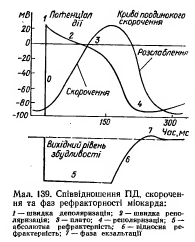
77.Изменения возбудимости во время развития ПД типичных кардиомиоцитов, их значение.
Значення великої тривалості ПД Т-КМЦ стає зрозумілим, якщо співставити його в часі з графіком зміни збудливості Т-КМЦ при збудженні з графіком поодинокого скорочення міокарда:
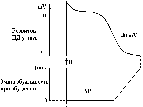
ПД Т-КМЦ тривалий через наявність фази плато.
Довготривалий ПД є причиною тривалої абсолютної рефрактерної фази (АР) – час протягом якого Т-КМЦ повністю незбудливі.
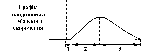
АР відповідає розвитку латентного періоду поодинокого м’язевого скорочення, періоду укорочення та значної частини періоду розслаблення.
1. Латентний період.
2. Період укорочення.
3. Період розслаблення.
Завдяки такому співвідношенню у часі фаз збудливості та періодів поодинокого скорочення міокарда досягається:
- неможливість виникнення в міокарді тетанічних скорочень; наступний цикл збудження (і скорочення) стає можливим тільки в фазі відносної рефрактерності, коли міокард закінчив своє скорочення і в значній мірі розслабився. Це дуже важливо, так як для виконання насосної функції серцем необхідно, щоб воно наповнилось кров’ю під час розслаблення. Тетанічне скорочення унеможливлювало б нагнітальну функцію серця;
- неможливість патологічної рециркуляції збудження по структурах серця (тривала фаза абсолютної рефрактерності не дає можливості збудженню повернутись туди, де воно було деякий час тому назад).
78.Современное представление о природе и градиенте автоматии сердца. Проводящая система сердца. Опыт Станниуса. Нарисовать ПД пейсмекерных клеток.
В
звичайних умовах послідовність руху
збудження по структурах серця така:
пазухово-передсердний вузол робочий
міокард передсердь передсердно-шлуночковий
вузол пучок Гіса ніжки пучка Гіса
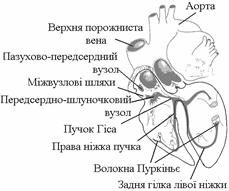
волокна Пуркіньє робочий міокард шлуночків.
Швидкість проведення збудження по структурах серця різна.
Вона складає:
- робочий міокард передсердь та шлуночків – біля 1 м/с;
- пучок Гіса, його ніжки, волокна Пуркіньє – 2-5 м/с.
Тобто, швидкість велика і це має велике значення, так як забезпечує синхронність скорочень міокарду передсердь; міокарду шлуночків (для досягнення останнього результату існує провідна ситема шлуночків серця – пучок Гіса, його ніжки, волокна Пуркіньє; наявність спеціальної провідної системи шлуночків зумовлено його великою масою).
Чинниками, що впливають на швидкість проведення збудження по м’язовим волокнам є: діаметр волокон, амплітуда ПД, величина порогу деполяризації, швидкість розвитку піку ПД, наявність нексусів між міокардіоцитами – вони мають низький опір, що сприяє швидкій передачі ПД з одного КМЦ на другий і збільшенню швидкості проведення збудження.
Причинами великої швидкості проведення збудження по провідній системі серця є:
- великий діаметр волокон;
- наявність нексусів;
- достатня амплітуда ПД (близько 100 мВ).
Дещо менший діаметр Т-КМЦ та гірше розвинені нексуси є причиною меншої швидкості проведення збудження по ним.
В ділянці передсердно-шлуночкового вузла має місце затримка проведення збудження, котра виникає внаслідок значного зменшення швидкості проведення (2-5 м/с). Наявність атріовентрикулярної затримки забезпечує послідовне скорочення передсердь та шлуночків (спочатку передсердя, а через 0,1 с – шлуночки).
Причини малої швидкості проведення збудження по волокнам передсердно-шлуночкового вузла є:
- малий діаметр волокон;
- відсутність нексусів;
- невелика амплітуда ПД (60-70 мВ);
- мала швидкість розвитку ПД (в цих клітинах розвиток ПД має не натрієву, а кальцієву природу, тому розвивається повільно).
Опыт
Станиусса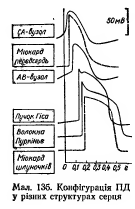
1-ю лигатуру накладывают между предсердиями и венозным синусом для изоляции последнего. Венозный синус продолжает сокращаться с прежней частотой, а предсердия и желудочек останавливаются. Водитель ритма сердца лягушки находится в венозном синусе.
2-ю лигатуру накладывают Между предсердиями и желудочком сердца для раздражения области атриовентрикулярного соединения. Желудочек возобновляет сокращения, но с меньшей частотой, чем венозный синус. В области атриовентрикулярного соединения имеется латентный (потенциальный) водитель ритма, или водитель ритма 2-го порядка.
3-ю лигатуру накладывают На уровне нижней трети желудочка с целью изоляции его верхушки. Последняя перестает сокращаться. В верхушке желудочка сердца лягушки нет водителя ритма.
Водитель ритма сердца лягушки находится в венозном синусе; имеется потенциальный (латентный) водитель ритма в области атриовентрикулярного соединения; верхушка желудочка сердца лягушки автоматией не обладает;существует убывающий градиент автоматии от основания сердца (области венозного синуса) к его верхушке.
79.ПД атипичных кардиомиоцитов сино-атриального узла, механизмы происхождения, физиологическая роль.
80.Электромеханическое сопряжение в миокарде. Механизм сокращения и расслабления миокарда.
Спряження (зв’язок) збудження і скорочення в міокарді принципово проходить так само, як і в скелетних м’язах. Тобто, ПД викликає скорочення таким чином:
ПД поширюється по мембрані Т-КМЦ, в тому числі і по мембрані Т-трубочок відкриття кальцієвих каналів саркоплазматичного ретикулума (СПР) вихід йонів кальцію із СПР підвищення концентрації йонів кальцію в міоплазмі з 10-8 до 10-5 моль/л дифузія йонів кальцію до скоротливих білків (протофібрил) взаємодія з регуляторними білками (з тропоніном) зміна третинної структури тропоніну та тропоміозину відкриття активних центрів актину взаємодія активних головок міозину з активними центрами актину скорочення міокарду.
Необхідно підкреслити, що сила серцевих скорочень (ССС) залежить від кількості актоміозинових містків, які утворюються при скороченні.
Особливостями процесу спряження збудження та скорочення в міокарді є:
- необхідність для виходу йонів кальцію із СПР (кальцієвого залпу) входу йонів кальцію із міжклітинної рідини. Цей вхід проходить під час фази плато ПД;
- наявність кількісного взаємозв’язку між входом кальцію в клітину під час фази плато ПД і його виходу із СПР, а отже, і ССС (регуляторні механізми, наприклад, можуть підвищувати кількість повільних кальцієвих каналів, через які проходить вхід йонів кальцію під час фази плато ПД підвищення входу кальцію під час фази плато ПД посилення виходу йонів Сa+ із СПР підвищення кількості відкритих активних центрів актину підвищення кількості акто-міозинових містків посилення ССС).
Стан кальцієвих каналів можуть змінювати лікарі, призначаючи хворим їх блокатори (варапаміл) зменшення входу йонів кальцію в Т-КМЦ при їх збудженні зменшення ССС.
Механізми розслаблення міокарду полягає у видаленні із міоплазми йонів кальцію, які надійшли туди під час “кальцієвого залпу”. Цей результат досягається завдяки:
- активації кальцієвих насосів повздовжніх трубочок СПР (із затратами енергії АТФ);
- активації кальцієвих насосів зовнішньої мембрани Т-КМЦ (із затратами енергії АТФ);
- роботі натрій-кальцієвого йонообмінного механізму; цей механізм забезпечує транспорт в протилежному напрямку через зовнішню клітинну мембрану йонів Na+ (в клітину) і йонів Сa+ (з клітини). Йони Na+ входять в клітину за градієнтом концентрації, який створюється натрій-калієвим насосом (працює з затратами енергії АТФ). Спряжений з входом йонів Na+ вихід йонів Сa+ знижує його концентрацію в клітині та сприяє розслабленню міокарда.
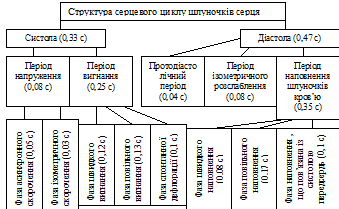
81.Сердечный цикл, его фазы, их физиологическая роль.
Серце в системі кровообігу виконує функцію насоса. Його будова повністю пристосована для виконання функцій насоса:
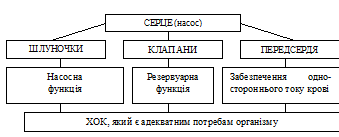
Таким чином, насосну функцію виконують, перш за все, шлуночки серця. Головна функція передсердь полягає в акумулюванні (накопиченні) крові при закритих передсердно-шлуночкових клапанах (кровообіг в судинах безупинний!).
Серце як насос працює циклічно – мають місце ритмічне чергування систоли (скорочення) та діастоли (розслаблення) відділів серця. В стані спокою ЧСС = 75 в хвилину, тривалість серцевого циклу (СЦ) складає 0,8 с. Чергування систоли та діастоли різних відділів серця можна представити у вигляді схеми (одна клітинка = 0,1 с):
Загальна
пауза – час протягом якого співпадає
діастола передсердь та шлуночків.
Основна функція передсердь – резервуарна, тому їх серцевий цикл простий та складається лише із систоли та діастоли.
Шлуночки виконують насосну функцію та мають складну структуру СЦ.
Перед детальною характеристикою окремих фаз та періодів СЦ варто підкреслити, що причиною руху крові через порожнини серця, із порожнин в судини і т.д., причиною зміни положення клапанів серця є градієнт тиску, який виникає внаслідок скорочення та розслаблення відділів серця. Тому події, що відбуваються в серці під час СЦ ми будемо розглядати в такій послідовності: скорочення (розслаблення) серця зміна тиску в його порожнинах виникнення градієнтів тисків, які зумовлюють:
а) зміну положення клапанів;
б) рух крові.
Характеристика періодів і фаз СЦ:
NB на прикладі ЛІВОГО СЕРЦЯ!!!
Початку нового СЦ передує загальна пауза. В її кінці тиск в шлуночку приблизно рівний 5 мм рт. ст., в передсерді він трішки вищий, а в венах тиск вищий, ніж в передсерді. При такому розподілі тиску мітральний клапан – відкритий; кров дуже повільно тече з передсердя в шлуночок, а із вен – в передсердя. Тиск в аорті вищий від діастолічного, тобто набагато вищий, ніж у шлуночку. Саме цей градієнт тиску тримає закритими півмісяцеві клапани.
СЦ починається з систоли передсердя. Її тривалість складає близько 0,1 с. Починається скорочення передсердя з м’язевих пучків, які охоплюють гирла вен; це попереджує рух крові по градієнту тиску із передсердя в вени, так як клапани тут відсутні. Тиск в передсерді в результаті його скорочення підвищується до 8 мм рт. ст. і внаслідок цього в шлуночок надходить остання порція крові, яка складає від 8% до 30% від всього об’єму крові, що надходить в шлуночок при його діастолі.
Вслід за систолою передсердя починається систола шлуночка, яка в загальному триває 0,33 с. Систола шлуночка складається з 2-ох періодів: