

37. Конечные продукты азотистого обмена: соли аммония и мочевина.
Наиболее значительные количества аммиака обезвреживаются в печени путём синтеза мочевины. В первой реакции процесса аммиак связывается с диоксидом углерода с образованием карбамоилфосфата, при этом затрачиваются 2 молекулы АТФ.
Мочевина - основной конечный продукт азотистого обмена, в составе которого из организма выделяется до 90% всего выводимого азота (рис. 9-15). Экскреция мочевины в норме составляет ~25 г/сут. При повышении количества потребляемых с пищей белков экскреция мочевины увеличивается. Мочевина синтезируется только в печени.
... Соли аммония - это сложные вещества, в состав которых входят ионы аммония NH4+, соединённые с кислотными остатками... УСЁ.
38. Основные источники и пути обезвреживания аммиака в организме.

Обезвреживание аммика может быть на месте, так где он собственно образуется, и возможны разные механизмы, такие как.
- Восстановительное аминирование α-кетоглутарата
- Амидирование аминокислот (синтез амидов)
- Амидирование карбоксильных групп белков
- Синтез карбамоилфосфата
- Синтез аммонийных солей
И конечно же, главный способ ликвидации это - синтез мочевины, которы расписан ниже.
39. Роль глутамина в обезвреживании и транспорте аммиака в организме.
Глутами - тот парень, который берет на себя всю основную нагрузку, связанный с работой с аммиаком. Основной реакцией связывания аммиака, протекающей во всех тканях организма, является синтез глутамина под действием глутамин-синтетазы:

Глутамин легко транспортируется через клеточные мембраны путём облегчённой диффузии (для глутамата возможен только активный транспорт) и поступает из тканей в кровь. Основными тканями-поставщиками глутамина служат мышцы, мозг и печень. С током крови глутамин транспортируется в кишечник и почки.
В клетках кишечника под действием фермента глутаминазы происходит гидролитическое освобождение амидного азота в виде аммиака. В почках тоже самое.
40. Глутамин как донор амидной группы при синтезе ряда соединений.
Глутамин - основной донор азота в организме.Амидный азот глутамина используется для синтеза пуриновых и пиримидиновых нук-леотидов, аспарагина, аминосахаров и других соединений

41. Синтез мочевины, химизм, ферменты, энергетика, происхождение атомов азота в мочевине.
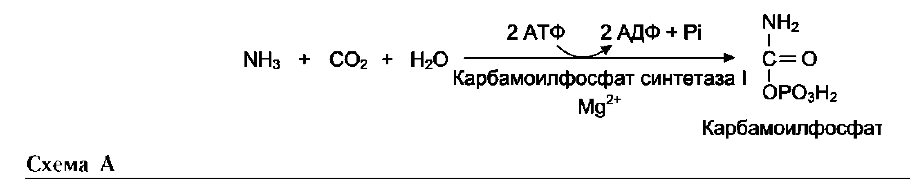
Мочевина (карбамид) - полный амид угольной кислоты - содержит 2 атома азота. Источником одного из них является аммиак, который в печени связывается с диоксидом углерода с образованием карбамоилфосфата под действием карбамоилфосфатсинтетазы I
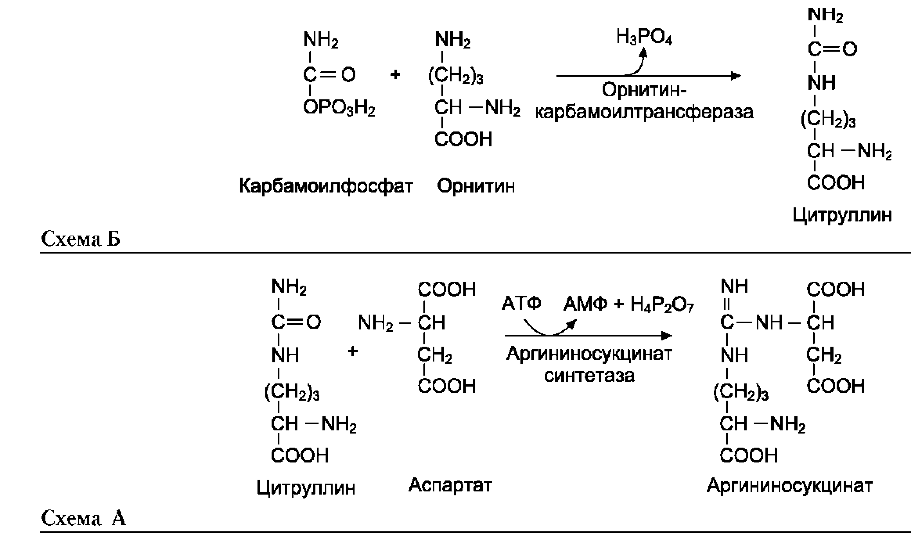
Далее под действием орнитин-карбамоил-трансферазы карбамоильная группа карбамо-илфосфата переносится на α-аминокислоту орнитин, и образуется другая α-аминокислота - цитруллин.
В следующей реакции аргининосукцинат-синтетаза связывает цитруллин с аспартатом и образует аргининосукцинат (аргининоянтарную кислоту). Этот фермент нуждается в ионах Mg2+. В реакции затрачивается 1 моль АТФ, но используется энергия двух макроэргических связей. Аспартат - источник второго атома азота мочевины
Далее фермент аргининосукцинатлиаза (арги-ниносукциназа) расщепляет аргининосукцинат на аргинин и фумарат, при этом аминогруппа аспартата оказывается в молекуле аргинина.
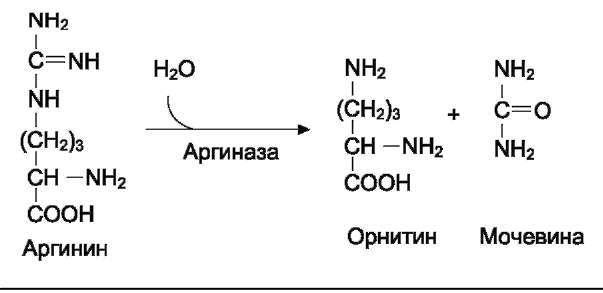
Аргинин подвергается гидролизу под действием аргиназы, при этом образуются орнитин и мочевина. Кофакторами аргиназы являются ионы Са2+ или Мп2+.


Всю эту красотень можно выразить формулой попроще.
СО2 + NH3 + Аспартат + 3 АТФ + 2 Н2О → Мочевина + Фумарат + 2 (АДФ + Н3РО4) + АМФ + Н4Р2О7
Энергетика ... загружаю... снятие грифа секретности...
В результате орнитинового цила расходуются четыре макроэргические связи трех молекул АТФ на каждый оборот цикла. Однако, все неплохо компенсируется:
- при включении фумарата в ЦТК на стадии дегидрирования малата образуется НАДН, который обеспечивает синтез трех молекул АТФ.
- при окислительном дезаминировании глутамата в разных органах также образуется НАДН, и образуется еще три молекулы АТФ.
Биологическая роль, две основные цели:
- превращение азота аминокислот в мочевину, которая экскретируется и предотвращает накопление токсичных продуктов, главным образом аммиака.
- синтез аргинина и пополнение его фонда в организме.
Основной источник аммиака - аминокислоты. Большая часть образовавшегося аммиака обезвреживается в орнитиновом цикле в печени и выделяется в виде мочевины. Основной реакцией обезвреживания аммиака в тканях является синтез глутамина, который затем используется в анаболических процессах и для обезвреживания веществ в печени.