

Свойства синапсов:
-
односторонность проведения возбуждения через синапс (всегда от пре- к постсинаптической мембране обусловлена наличием рецепторов чувствительных к медиатору только на постсинаптической мембране;
-
синаптическая задержка - замедление скорости распространения возбуждения в синапсе в связи с длительностью процессов выделения медиатора из пресинаптического окончания, диффузии его по синаптической щели и процесса взаимодействия его с постсинаптической мембраной. Синаптическая задержка в химических синапсах обычно равна 0,2 - 0,5 мс.
-
низкая лабильность - обусловлена временем распространения предыдущего импульса и наличием у него периода абсолютной рефрактерности;
-
повышенная утомляемость - обусловлена отставанием скорости синтеза медиатора от скорости его распада при возбуждении;
-
трансформация ритма возбуждения, обусловлена тем, что не все приходящие к синаптическому окончанию аксона потенциалы действия способны вызвать критическую деполяризацию постсинаптической мембраны;
-
высокая чувствительность к лекарствам и ядам обусловлена высокой специфичностью хеморецепторов постсинаптической мембраны;
-
низкий фактор безопасности (надежности), обусловлен тем, что хеморецепторы чувствительны к большому количеству веществ, способных инициировать или изменять активность синапса.
Адренергичеcкие синапсы
Адренергические нейроны расположены в ЦНС (голубое пятно среднего мозга, мост, продолговатый мозг) и в симпатических ганглиях.
Периферические адренергические синапсы образованы варикозными утолщениями разветвлений постганглионарных симпатических волокон.
Медиатор адренергических синапсов - норадреналин. Его предшественник в биосинтезе дофамин выполняет медиаторную функцию в дофаминергических синапсах. Адреналин представляет собой гормон мозгового слоя надпочечников. Все три вещества относятся к группе катехоламинов, так как содержат гидроксилы в 3-м и 4-м положениях ароматического кольца.
Синаптические пузырьки в адренергических синапсах имеют под электронным микроскопом гранулярное строение и поэтому получили название гранулы.
В гранулах норадреналин депонирован в связи с АТФ и белком хромогранином. В составе гранул обнаружены также ферменты и модулирующие нейропептиды (энкефапины, нейропептид Y).
Норадреналин синтезируется из аминокислоты тирозина. Превращение фенилаланина в тирозин является неспецифическим процессом и происходит в печени. Обе аминокислоты в большом количестве содержатся в твороге, сыре, шоколаде, бобовых.
ТИРОЗИН
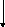 Тирозингидроксилаза
Тирозингидроксилаза
(тирозин-3-монооксигеназа)
Кофактор - тетрагидроптерин
ДОФА
Д екарбоксилаза
ароматических
екарбоксилаза
ароматических
L- аминокиспот
Кофактор - пиридоксальфосфат
Д ОФАМИН
ОФАМИН
Дофамин-β-гидроксилаза
Кофактор – аскорбат
Н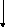 ОРАДРЕНАЛИН
ОРАДРЕНАЛИН
Фенилэтаноламин-N-метилтрансфераза Кофактор – S – аденозилметионин
АДРЕНАЛИН
После диссоциации комплексов норадреналин - адренорецептор медиатор инактивируется при участии ряда механизмов:
• Нейрональный захват (захват-1) - активный транспорт вначале через пресинаптическую мембрану (сопряжен с выходом ионов натрия), а затем через мембрану гранул под влиянием АТФ-зависимой протонной транслоказы (при входе в гранулы одной молекулы норадреналина в цитоплазму выходят 2 протона);
• Экстранейрональный захват (захват-2) нейроглией, фибробластами, миокардом, эндотелием и гладкими мышцами сосудов;
• Инактивация ферментами.
70-80% норадреналина участвует в нейрональном захвате, по 10% подвергается экстранейрональному захвату и ферментативному расщеплению. Необходимость нейронального захвата диктуется дефицитом субстратов и большой потребностью в энергии для синтеза норадреналина из тирозина. Для сохранения адреналина основное значение имеет экстранейрональный захват.
Ферменты инактивации катехоламинов - моноаминоксидаза (МАО) и катехол – О -метилтрансфераза (КОМТ). МАО, локализованная на внешней мембране митохондрий и в гранулах, осуществляет окислительное дезаминирование катехоламинов с образованием биогенных альдегидов. Затем альдегиды окисляются НАД-зависимой альдегиддегид-рогеназой в кислоты или восстанавливаются альдегидредуктазой в гликоли.
Цитоплазматический фермент КОМТ катализирует присоединение метильной группы к гидроксилу в 3-м положении ароматического кольца (только при наличии гидроксила в 4-м положении). Донатором метильных групп служит S-аденозилметионин. Метилированные продукты в 200-2000 раз (по разным тестам) менее активны, чем норадреналин и адреналин.