

Part five. Development of the spinal cord and brainstem nuclei
The cranial end of the neural tube (see p. 23) becomes dilated into vesicles and its walls thicken by proliferation of cells; the cerebral hemispheres, brainstem and cerebellum are so developed. More caudally the neural tube enlarges in a simple manner by proliferation of cells, to form the spinal cord. In all regions these proliferating cells arrange themselves regularly in functional groups. Despite the apparent random arrangement of the nuclei in the brainstem, there is a logical pattern to their positions which can be correlated with the rather simpler disposition of cell groups in the spinal cord that results from this development.
Spinal cord
The central canal, relatively very large at first, is not rounded in cross-section, but is projected laterally into a groove on the inner wall of the spinal cord, as the sulcus limitans (Fig. 7.33A). The developing nerve cells of the spinal cord form thickenings dorsal and ventral to the sulcus, the alar lamina and basal lamina respectively. The alar lamina contains sensory (afferent) cells and the basal lamina contains motor (efferent) cells. In each lamina the cells are of two kinds; near the sulcus limitans lie the autonomic (visceral) cells, while further away lie the body wall and limb (somatic) cells.
Figure 7.33 Cell groups of the spinal cord and brainstem: A in the developing spinal cord, where motor cells are ventral and sensory cells dorsal with autonomic cells in between; B in the brainstem (floor of the fourth ventricle) which resembles an ‘opened out’ spinal cord, with motor cells now lying medially and sensory cells laterally with autonomic cells in between.
Brainstem
A similar arrangement holds in the brainstem as in the spinal cord. But here a third type of cell appears in each lamina, namely the branchial afferent and efferent cells of cranial nerves supplying the derivatives of the branchial arches (see p. 24). These branchial cells theoretically lie between the autonomic and somatic cells of each lamina. They are the central cell stations of the nerves of the pharyngeal arches (trigeminal, facial, glossopharyngeal and vagus). The branchial group or column is also known as the special visceral group, and the visceral as the general visceral group.
In the fourth ventricle the central canal is opened out, and basal and alar laminae lie roughly in the same plane; the dorsal afferent and ventral efferent cells thus become lateral and medial respectively (Fig. 7.33B). The order of the cell groups is similar, but migration of certain cell groups (neurobiotaxis) in the developing brainstem alters, in places, the relatively simple basic arrangement.
Motor nuclei of the brainstem
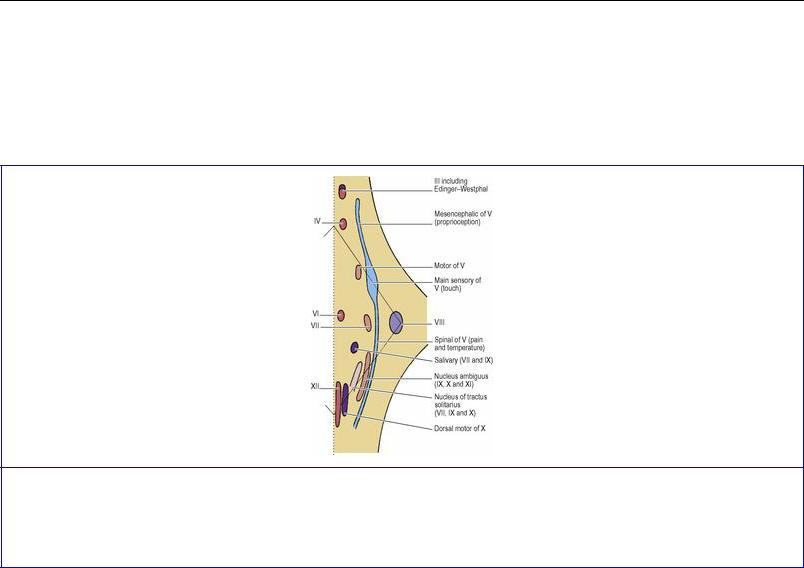
The motor nuclei are arranged according to the type of muscle they supply (Fig. 7.34). The ordinary skeletal muscle of the head (somatic muscle) consists of the muscles of the orbit and the muscles of the tongue. In line with the anterior horn cells of the spinal cord, the brainstem nuclei supplying these muscles (oculomotor, trochlear, abducent and hypoglossal) lie near the midline ventral to the ‘central canal’ (i.e. ventral to the aqueduct or floor of the fourth ventricle as the case may be).
Figure 7.34 Sites of cranial nerve nuclei in the right half of the brainstem as seen from the dorsal aspect. See text for the correlation between the information in this diagram with that in Figure 7.33B.
Developed from the region of the embryonic pharynx are the striated muscles of mastication, of the face, and of the pharynx and larynx. Their motor nuclei (branchial) lie slightly more laterally: the motor nucleus of the trigeminal nerve, the facial nucleus and the nucleus ambiguus (for the glossopharyngeal and vagus nerves and the cranial part of the accessory nerve).
T he general visceral efferent, parasympathetic motor, nuclei are represented by the accessory oculomotor (Edinger–Westphal) nucleus, the salivary nucleus (whose secretomotor fibres join the nervus intermedius part of the facial nerve and the glossopharyngeal nerve), and the dorsal motor nucleus of the vagus (for cardiac muscle and the smooth muscle of the alimentary tract). These nuclei migrate medially.

Afferent nuclei of the brainstem
These follow the general plan outlined above, and from medial to lateral form visceral, branchial and somatic afferent groups (the reverse order of the motor efferent columns). But unlike the motor groups, which consist of interrupted columns forming individual nuclei, each of these afferent ‘columns’ consists of a single nuclear mass ( Fig. 7.34). The general visceral and branchial (special visceral) afferent cell groups merge as the nucleus of the tractus solitarius, which receives not only taste fibres from the facial, glossopharyngeal and vagus nerves but also afferents from the heart, lungs and other viscera from the vagus nerve. The somatic afferent column is represented by the sensory nuclei of the trigeminal nerve, really one elongated nucleus extending throughout the brainstem to the upper spinal cord. The brainstem has a fourth column of sensory nuclei that has no counterpart in the spinal cord; these are the cochlear and vestibular nuclei of the vestibulocochlear nerve, and they form a special somatic afferent group lying farthest laterally.
Part six. Summary of cranial nerves
The first two cranial nerves are not really nerves, but rather outdrawn parts of the central nervous system. Only the peripheral bipolar nerve cells correspond with an ordinary nerve.
Olfactory nerve (i)
The first neurons are bipolar cells in the neuroepithelium in the upper part of the nose. Their central processes gather themselves into about 20 olfactory nerve filaments (for each olfactory nerve) that pass through foramina in the cribriform plate of the ethmoid bone, piercing the dura and arachnoid mater of the anterior cranial fossa. They enter the olfactory bulb from where the central processes of the second neuron pass in the olfactory tract to the region of the anterior perforated substance, from where they proceed laterally to the uncus and medially to parts of the limbic system. The arrangement is unique and ‘primitive’: the second order neuron directly activates the conscious cortex, bypassing the thalamus. Other olfactory pathways, by polysynaptic junctions, activate hypothalamic and brainstem nuclei (as is the case with all sensory pathways) for visceral and somatic effects, distinct from conscious appreciation.
Optic nerve (ii)
The rods and cones, near the choroidal surface of the retina, activate the bipolar cells of the retina; these are the first sensory neurons. Their central processes synapse on the large ganglion cells that lie on the vitreous surface of the retina. The central processes of these second neurons emerge from the back of the eyeball as the optic nerve surrounded by CSF and the meninges. The optic nerve enters the middle cranial fossa through the optic canal and passes to the chiasma, where the nasal fibres from each retina decussate, above the pituitary gland. From the chiasma the optic tract passes around the midbrain (cerebral peduncle) to three destinations: (1) the lateral geniculate body (thalamus) for relay to the visual cortex; (2) the pretectal nuclei for pupil constriction to light; and (3) the superior colliculus for body reflexes to light.
From the lateral geniculate body fibres of the third neurons of the visual pathway pass through the retrolentiform part of the internal capsule and backwards by the optic radiation to the visual (striate) cortex on the medial surface of the occipital lobe.
Oculomotor nerve (iii)
The nerve emerges from the midbrain on the medial surface of the base of the cerebral peduncle above the pons. It passes forwards and laterally between the posterior cerebral and superior cerebellar branches of the basilar artery, then below and lateral to the posterior communicating artery and just below the free margin of the tentorium cerebelli. It enters the roof of the cavernous sinus and continues forward in the lateral wall of the sinus. It slants down medial to the trochlear nerve and the ophthalmic branch of the trigeminal (see Fig. 6.104, p. 446). At the anterior pole of the cavernous sinus it splits into a superior and an inferior division which enter the tendinous ring at the medial end of the superior orbital fissure.
The superior division supplies superior rectus and levator palpebrae superioris. While in the lateral wall of the cavernous sinus, the oculomotor nerve picks up sympathetic fibres from the internal carotid plexus; they supply the smooth muscle part of levator (their loss accounts for the partial ptosis characteristic of Horner's syndrome; see p. 408).
T h e inferior division supplies medial rectus, inferior rectus and inferior oblique. The parasympathetic fibres in the oculomotor nerve pass from the branch to the inferior oblique to the ciliary ganglion, where they relay. Postganglionic fibres then run in the short ciliary nerves to supply the sphincter pupillae (for pupil constriction) and the ciliary muscle (for accommodation).
Trochlear nerve (iv)
The nerve crosses the midline within the midbrain and emerges from its dorsal aspect below the inferior colliculus. It then passes around the cerebral peduncle between the posterior cerebral and superior cerebellar arteries. Clinging to the undersurface of the free edge of the tentorium cerebelli it is directed thereby to the roof of the cavernous sinus, which it enters and then runs forwards in the lateral wall, first below the third nerve, and then above it at the anterior end of the sinus. It enters the superior orbital fissure lateral to the tendinous ring and passes medially over levator palpebrae superioris to supply superior oblique.
Trigeminal nerve (v)
The fibres of the trigeminal nerve that reach the main sensory and spinal nuclei come from cell bodies in the trigeminal ganglion. Together with the direct fibres that reach the mesencephalic nucleus they form a single, large, sensory root attached to the ventral aspect of the pons, well lateral and just above centre. The motor root emerges separately, slightly cranial and medial to its companion. Together they pass, below the tentorium cerebelli, to the mouth of the trigeminal cave (see Fig. 6.105, p. 446). This is a tubular prolongation of arachnoid-lined fibrous dura mater around the sensory and motor roots, and it crosses the upper border of the petrous bone near its apex, i.e. it passes from the posterior into the middle cranial fossa. The dural sheath containing the two nerve roots passes forwards, peeling apart the two layers of dura that floor the middle cranial fossa, just lateral to where the same two layers peel apart to enclose the cavernous sinus. The sensory root then expands into the large, flat, crescentic trigeminal ganglion; the motor root remains separate below the ganglion. The dural sheath obliterates the subarachnoid space by fusing with the pia mater halfway along the ganglion. The posterior half of the ganglion and both roots are thus bathed in CSF. The anterior half of the ganglion, beyond the subarachnoid space, gives off its three sensory divisions, ophthalmic, maxillary and mandibular. The first two pass forwards in the lateral wall of the cavernous sinus; they are wholly sensory. The mandibular division, likewise sensory, passes straight down from the lower part of the ganglion to the foramen ovale. The motor root also passes through the foramen ovale and joins the mandibular division just outside the skull to form the (mixed) mandibular nerve.
Ophthalmic nerve
As the ophthalmic division runs forward in the lateral wall of the cavernous sinus below the trochlear nerve it picks up sympathetic fibres from the plexus around the internal carotid artery; these are for the dilator pupillae muscle. At the anterior end of the sinus it gives off a meningeal branch and divides into three branches that pass through the superior orbital fissure: the lacrimal; frontal; and nasociliary nerves.
The lacrimal nerve, passing lateral to the tendinous ring, proceeds along the upper part of the lateral wall of the orbit, there picking up a secretomotor branch from the zygomaticotemporal branch of the zygomatic nerve which it gives to the lacrimal gland. It is sensory to a small area of skin at the lateral end of the upper eyelid and to both palpebral and ocular surfaces of the corresponding conjunctiva.
The frontal nerve traverses the superior orbital fissure just lateral to the tendinous ring. A large nerve, it runs forward above levator palpebrae superioris, and behind the superior orbital margin it divides into a large supraorbital and a small supratrochlear branch. The supraorbital nerve supplies the frontal sinus, notches or perforates the orbital margin, supplies the upper eyelid (skin and both surfaces of conjunctiva), all the forehead except a central strip, and the frontal scalp up to the vertex. The supratrochlear nerve supplies the upper lid and conjunctiva and a narrow strip of forehead skin alongside the midline.
The nasociliary nerve is sensory to the whole eyeball, to the paranasal sinuses along the medial wall of the orbit, to some mucous membrane of the nasal cavity and to the skin of the external nose. It carries hitch-hiking sympathetic fibres for the dilator pupillae muscle.
The nerve runs through the tendinous ring between the two divisions of the oculomotor nerve. It passes forwards and medially above the optic nerve, below superior rectus and superior oblique. At the medial wall of the orbit it passes into the anterior ethmoidal foramen, between the orbital plate of the ethmoid bone and the frontal bone, as the anterior ethmoidal nerve. Running in the roof of the middle and anterior ethmoidal air cells, it reaches the lateral edge of the cribriform plate, beneath the dura of the anterior cranial fossa. It then runs forwards and descends through a slit alongside the crista galli into the nose, where it grooves the internal surface of the nasal bone. It supplies the mucous membrane of the anterosuperior part of the lateral wall and septum of the nose, and emerges at the lower margin of the nasal bone as the external nasal nerve, to supply the skin of the ala, tip and vestibule of the nose.
The infratrochlear nerve branches off the nasociliary before it the enters the anterior ethmoidal foramen, and continues forward below the trochlea of the superior oblique tendon to supply skin and conjunctiva of the medial end of the upper eyelid and skin over the bridge of the nose.
The posterior ethmoidal nerve branches off the nasociliary proximal to the infratrochlear nerve, enters the posterior ethmoidal foramen and supplies the posterior ethmoidal air cells and the adjacent sphenoidal sinus.
In the orbit the nasociliary nerve gives off a communicating branch to the ciliary ganglion and the long ciliary nerves. The communicating branch is the sensory root of the ganglion; its fibres pass through the ganglion and via the short ciliary nerves to provide sensory fibres to the eye including the cornea
(but not the conjunctiva). The long ciliary nerves, usually two, run forward to enter the sclera independently. They carry sympathetic fibres to the dilator pupillae muscle and (like the short ciliary nerves) are also sensory to the eye, including the cornea (but not the conjunctiva).

Maxillary nerve
This nerve runs forward in the lateral wall of the cavernous sinus, below the ophthalmic nerve, gives off a meningeal branch and passes through the foramen rotundum into the upper part of the pterygopalatine fossa. It has a short course in the fossa, and turns laterally as it passes through the inferior orbital fissure to enter the orbit as the infraorbital nerve.
Two ganglionic branches connect the maxillary nerve and the pterygopalatine ganglion. Most of the fibres of the trunk pass through the ganglion into its branches, where they mingle with the postganglionic fibres of the greater petrosal and deep petrosal nerves (i.e. fibres of the nerve of the pterygoid canal; see p. 370). The branches of the ganglion are nasal (nasopalatine and posterior superior nasal), palatine (greater and lesser), pharyngeal and orbital. Postganglionic parasympathetic fibres destined for the lacrimal gland pass into the maxillary nerve and enter its zygomatic branch.
The nasopalatine nerve (long sphenopalatine) enters the sphenopalatine foramen, crosses the roof of the nose and slopes down along the nasal septum, supplying its posteroinferior part. It goes through the incisive canal and fossa into the hard palate and supplies the gum behind the two incisor teeth.
The posterior superior nasal nerves (lateral and medial) enter the sphenopalatine foramen and supply the posterosuperior part of the lateral wall of the nose and nasal septum respectively.
The greater palatine nerve runs down in the greater palatine canal, between the perpendicular plate of the palatine bone and the body of the maxilla. Multiple branches supply the posteroinferior part of the lateral wall of the nose and the adjacent floor of the nose. The nerve emerges from the greater palatine foramen and supplies all the hard palate except the incisor gum.
The lesser palatine nerves also descend through the greater palatine canal but emerge from the lesser palatine foramina in the palatine bone and pass back to supply the mucous membrane on both surfaces of the soft palate and on the tonsil.
The pharyngeal nerve passes back through the palatovaginal canal to supply the mucous membrane of the nasopharynx.
The zygomatic nerve arises from the maxillary in the pterygopalatine fossa and passes through the inferior orbital fissure into the orbit, where it divides into two branches. The zygomaticofacial nerve perforates the facial surface of the zygomatic bone and supplies the skin over the bone. The zygomaticotemporal nerve perforates the temporal surface of the zygomatic bone, pierces the temporalis fascia, and supplies skin above the zygomatic arch. Through a connection between this nerve and the lacrimal nerve in the orbit secretomotor fibres are transmitted to the lacrimal gland.
The posterior superior alveolar nerve (posterior superior dental) arises from the maxillary in the pterygopalatine fossa and divides into two or three nerves which emerge through the pterygomaxillary fissure. They run in or on the posterior wall of the maxilla to supply the maxillary sinus, upper molar teeth and the adjacent gum of the vestibule.
The infraorbital nerve passes forward along the floor of the orbit, sinks into a groove, then enters a canal and emerges on the face through the infraorbital foramen. It supplies the maxillary sinus directly and via the middle superior alveolar and anterior superior alveolar nerves. The former nerve
supplies the premolar teeth and the latter nerve supplies the canine and the two incisors, and reaches the anterior inferior part of the lateral wall and the adjacent floor of the nose. It ends on the nasal septum. All the alveolar nerves link up to form a dental plexus; the middle nerve may be absent.
Emerging on the face the infraorbital nerve lies between levator labii superioris and levator anguli oris. It has many communications with branches of the facial nerve. It supplies both surfaces of the lower lid conjunctiva, as well as the skin of the lower lid, mid-face, nose and the skin and mucous membrane of the whole upper lip and adjacent gum.

Mandibular nerve
The mandibular division passes down through the foramen ovale, accompanied by the small motor root of the trigeminal nerve. Just below the foramen they join to form the mixed nerve in the infratemporal fossa, between the upper head of the lateral pterygoid and tensor palati, where the otic ganglion lies on the medial aspect of the nerve. The nerve soon divides into a small anterior and a large posterior branch.
There are two branches from the short trunk before it divides. A meningeal branch (nervus spinosus) passes up through the foramen ovale or through the foramen spinosum.
The nerve to the medial pterygoid sinks into the deep surface of the muscle. It has a branch that passes through the otic ganglion and supplies the two tensor muscles, tensor palati and tensor tympani.
Anterior division. The branches are all motor except one. They are as follows.
The nerve to the lateral pterygoid; this may be double, one to each head.
Two deep temporal nerves pass above the upper head of the lateral pterygoid and sink into the deep surface of temporalis.
The nerve to masseter passes above the upper head of the lateral pterygoid and proceeds laterally through the mandibular notch to sink into masseter. It gives a branch to the mandibular joint.
The buccal nerve is the only sensory branch of the anterior division. It emerges between the two heads of the lateral pterygoid and passes down on the lower head to reach the buccinator. It supplies a small area of cheek skin and pierces the buccinator to supply the mucous membrane adherent to the deep surface of the muscle and the vestibular gum of the three mandibular molar teeth. The buccal nerve carries secretomotor fibres from the otic ganglion for mucous glands in the mouth.
Posterior division. The three branches are all sensory except for motor fibres in the inferior alveolar nerve, which pass to its mylohyoid branch.
The auriculotemporal nerve has two roots that pass back around the middle meningeal artery. The nerve picks up postganglionic secretomotor fibres from the otic ganglion for the parotid gland. It passes back deep to the neck of the mandible, and gives the major sensory supply to the mandibular joint. It then ascends behind the joint and the superficial temporal vessels, in front of the ear and in contact with the parotid gland to which it gives the secretomotor fibres. The nerve supplies the external acoustic meatus, the external surface of the auricle above this and the skin of the temporal region.
The inferior alveolar nerve (inferior dental) passes down deep to the lower head of the lateral pterygoid, on the lateral surface of the medial pterygoid. Lying between the mandible and the sphenomandibular ligament, it enters the mandibular foramen in front of the inferior alveolar artery and vein, after giving off the nerve to mylohyoid. This pierces the sphenomandibular ligament and lies on the mylohyoid groove with its accompanying small vessels. The mylohyoid nerve passes forwards below the mylohyoid muscle accompanied by the submental branches of the facial artery and vein. It supplies mylohyoid and the anterior belly of the digastric, and frequently a small area of
submental skin.
The inferior alveolar nerve runs forwards in the mandibular canal and supplies the three molars and two premolars. Then it divides into its two terminal branches. The incisive branch goes on to supply the canine and both incisors, and overlaps to the opposite central incisor. The mental nerve passes from the mental foramen to supply the lower lip (both surfaces) and the adjacent gum. It carries a few fibres from the otic ganglion to the labial glands of the lower lip.
The lingual nerve is joined by the chorda tympani (carrying nervus intermedius fibres) about 2 cm below the base of the skull, deep to the lower border of the lateral pterygoid muscle. It curves down on the medial pterygoid in front of the inferior alveolar nerve. It then passes under the free lower border of the superior constrictor and goes forward above the mylohyoid muscle (i.e. in the mouth). It grooves the thin lingual plate of the mandible just below the last molar tooth, and here the nerve is characteristically flat. Dipping down lateral to the submandibular duct, the nerve turns medially below the duct and ascends on hyoglossus to the anterior two-thirds of the tongue, which it supplies with common sensation and taste, the latter mediated by the chorda tympani fibres. The secretomotor fibres of the chorda tympani are given off to the submandibular ganglion, which is suspended from the lingual nerve; they relay in the ganglion for the submandibular gland, and some postganglionic fibres rejoin the lingual nerve for transport to the salivary glands in the floor of the mouth. The lingual nerve supplies all the mucous membrane of the floor of the mouth and the lingual gum.
Abducens nerve (vi)
The nerve emerges at the lower border of the pons, above the pyramid of the medulla. It enters the pontine cistern and turns upwards, between the anterior inferior cerebellar artery and the pons, to pierce the arachnoid and dura mater on the clivus. Running up now between the two layers of the dura on its own, or within the inferior petrosal sinus, it passes over the apex of the petrous temporal bone to enter the cavernous sinus. It passes forwards in the sinus, inferolateral to the internal carotid artery, and reaches the medial end of the superior orbital fissure. It enters the orbit through the tendinous ring to supply lateral rectus.
Facial nerve (vii)
The main facial nerve emerges at the lower border of the pons above the olive.
The nervus intermedius emerges between the pons and the inferior cerebellar peduncle, near the vestibulocochlear nerve. With the main part of the facial nerve it passes laterally in the cerebellopontine angle through the pontine cistern and, with the eighth nerve, they enter the internal acoustic meatus. In the meatus the main part of the facial nerve lies on the upper surface of the eighth nerve, with the nervus intermedius in between. The whole facial nerve passes into the anterosuperior quadrant of the fundus of the meatus. Running laterally in the petrous bone, above the vestibule of the internal ear, the two parts of the nerve share a common tube of arachnoid and dura mater. The meninges then fuse with the nerves as they merge into a single trunk. Near the middle ear the nerve makes a sharp posterior bend, the geniculum, which is enlarged by the cell bodies of the afferent (taste) fibres to form the geniculate ganglion. From here the nerve runs back in the medial wall of the middle ear, above the promontory and just below the bulge of the lateral semicircular canal. It now curves downwards behind the middle ear, deep to the aditus to the antrum, and passes vertically down the facial canal. After shedding all the nervus intermedius fibres, the nerve emerges from the stylomastoid foramen as a purely motor nerve and passes through the parotid gland. The face muscles supplied by the facial nerve receive their proprioceptive supply from the cutaneous nerves of the overlying skin (trigeminal nerve branches).
The intracranial branches of the facial nerve that arise within the petrous bone include the greater petrosal nerve, the nerve to stapedius and the chorda tympani.
The greater petrosal nerve, consisting of nervus intermedius fibres, leaves the ganglion and travels forwards and medially at a 45° slant through the petrous bone. It emerges from the anterosuperior surface of the petrous bone and runs forwards in a groove on the bone, between the two layers of the dura mater. Here in the middle cranial fossa it may be pulled on in extradural operations and so cause a small haemorrhage or oedema at the geniculate ganglion with consequent pressure on the facial nerve and a temporary facial paresis. The nerve passes beneath the trigeminal ganglion and reaches the foramen lacerum, where it is joined by the deep petrosal nerve from the sympathetic plexus on the internal carotid artery. The two unite and pass forwards through the pterygoid canal. This nerve of the pterygoid canal emerges into the pterygopalatine fossa and enters the pterygopalatine ganglion. Here the secretomotor fibres relay. Taste fibres from the soft palate and sympathetic fibres pass straight through the ganglion. Postganglionic secretomotor fibres are distributed with the branches of the ganglion to the nose, paranasal sinuses, hard and soft palates, and nasopharynx. Lacrimatory postganglionic fibres join the maxillary nerve and enter the orbit in its zygomatic branch.
T he nerve to stapedius is given off in the facial canal and reaches the muscle by a minute canaliculus.
The chorda tympani, consisting of nervus intermedius fibres, leaves the facial nerve in the facial canal 6 mm above the stylomastoid foramen and passes through the posterior wall of the middle ear. It runs forward between the mucous membrane and the tympanic membrane, crossing the neck of the malleus. It leaves through the anterior wall of the middle ear and emerges at the medial end of the petrotympanic fissure. It then grooves the medial side of the spine of the sphenoid and slopes downwards and forwards to join the lingual nerve in the infratemporal fossa. By the lingual nerve its
taste fibres are taken to the anterior part of the tongue. Its secretomotor fibres relay in the submandibular ganglion for the submandibular gland and glands in the floor of the mouth cavity.
T h e extracranial branches include the posterior auricular nerve (for the occipital belly of occipitofrontalis), the nerves to the posterior belly of digastric and stylohyoid, and the five groups of branches given off within the parotid gland for the facial muscles and platysma.
Vestibulocochlear nerve (viii)
Cochlear nerve
The receptors for hearing are the hair cells of the spiral organ in the internal ear. The cell bodies of the first neurons are in the spiral ganglion, in the base of the bony spiral lamina. Their central processes run along the modiolus of the cochlea and join into many small nerves that pierce dura and arachnoid mater at the base of the modiolus in a spiral pattern, at the anteroinferior quadrant of the fundus of the internal acoustic meatus. They join together in the subarachnoid space and enter the pontine cistern combined with the vestibular part. Together with the nervus intermedius and the main facial nerve, the vestibulocochlear nerve passes through the cerebellopontine angle in front of the flocculus of the cerebellum and the lateral aperture of the fourth ventricle. The eighth nerve enters the inferior cerebellar peduncle at the lower border of the pons.

Vestibular nerve
The receptors consist of the hair cells in the maculae of utricle and saccule (for static balance) and the ampullae of the semicircular ducts (for kinetic balance). From the posterosuperior quadrant of the fundus of the internal acoustic meatus emerges the superior division of the vestibular nerve from anterior and lateral semicircular ducts and the utricle. Through the posteroinferior quadrant comes the inferior division of the vestibular nerve from the saccule. Alongside it, through the foramen singulare, its branch emerges from the posterior semicircular duct. Having pierced the fibrous dura and arachnoid mater the upper and lower divisions lie in the internal acoustic meatus and are here together distended into the vestibular ganglion by the cell bodies of these first order neurons. From the ganglion the vestibular nerve joins the cochlear nerve and passes through the pontine cistern.
Glossopharyngeal nerve (ix)
The nerve emerges from the surface of the medulla between olive and inferior cerebellar peduncle in a series of rootlets which join to make a single nerve that runs laterally in the pontine cistern and enters the anterior compartment of the jugular foramen. Here it lies lateral to the inferior petrosal sinus, and together they are separated from the vagus and accessory nerves by a septum of fibrous dura mater. The glossopharyngeal nerve has a small superior ganglion and a large inferior ganglion which deeply notches the inferior border of the petrous bone, just below the internal acoustic meatus. The cell bodies of the nerve's afferent fibres are in these ganglia.
The tympanic branch passes into the middle ear through the tympanic canaliculus, between the jugular fossa and the carotid canal, to form the tympanic plexus, from which emerges the lesser petrosal nerve, on the floor of the middle cranial fossa. The lesser petrosal nerve runs through the foramen ovale to join the otic ganglion for the supply of the parotid gland.
The glossopharyngeal nerve passes laterally between the internal jugular vein and internal carotid artery and then passes forwards between the latter and the external carotid artery. The nerve to stylopharyngeus, given off as the nerve winds round the muscle, is the only muscular branch.
The carotid branch runs down to innervate the carotid sinus and body.
Pharyngeal branches take part (with the vagus) in forming the pharyngeal plexus; the fibres are afferent.
The glossopharyngeal nerve enters the pharynx between the superior and middle constrictors. The tonsillar branch provides afferent fibres for the tonsillar mucosa, and the lingual branch conveys common sensation and taste from the posterior part of the tongue, as well as secretomotor fibres for lingual glands.
Vagus nerve (x)
Vagal nerve fibres leave the surface of the medulla in a series of rootlets below those of the glossopharyngeal nerve in the sulcus between olive and inferior cerebellar peduncle. These unite into a single nerve that enters the middle compartment of the jugular foramen, with the accessory nerve, sharing an arachnoid and dural sheath. The vagus has a small superior ganglion just above the long inferior ganglion, which lies in the jugular fossa below the skull base. The superior ganglion has cell bodies for the auricular branch. The inferior ganglion lodges the cell bodies of all the other sensory fibres in the vagus nerve. The fibres of the cranial root of the accessory nerve join the vagus just above the inferior ganglion and the accessory nerve thereby gives its nucleus ambiguus fibres to the vagus.
In the neck the vagus lies vertical, like a plumb line, in the carotid sheath, deep in the gutter between the internal/ common carotid artery and the internal jugular vein.
The small meningeal, auricular and carotid body branches are all afferent. The meningeal branch carries C1 and C2 fibres which join the vagus at its exit from the skull.
The pharyngeal branch passes between the internal and external carotid arteries and provides both motor and sensory fibres for the pharyngeal plexus. The superior laryngeal branch runs down deep to the carotids and divides into the internal laryngeal nerve, which pierces the thyrohyoid membrane to supply mucosa in the pharynx and larynx, and the external laryngeal nerve which runs to cricothyroid. Cervical cardiac branches, upper and lower on the right and upper on the left, join the deep part of the cardiac plexus; the lower one on the left joins the superficial part of the plexus.
The right recurrent laryngeal nerve hooks under the subclavian artery at the root of the neck and runs up near the tracheo-oesophageal border to pass under the inferior constrictor of the pharynx just behind the cricothyroid joint, to supply muscles of the larynx and laryngeal mucosa of the vocal folds and below; it also supplies cricopharyngeus. The left recurrent laryngeal nerve hooks round the ligamentum arteriosum, below the aortic arch and ascends in a groove between the trachea and oesophagus to a corresponding destination. Both recurrent laryngeal nerves give cardiac branches.
In the superior mediastinum, the left vagus lies between the left common carotid and subclavian arteries, and not in contact with the trachea; it then crosses the left side of the aortic arch. The right vagus comes into contact with the trachea, posterior to the superior vena cava. Both vagi pass behind the lung roots, contributing branches to the anterior and posterior pulmonary plexuses. Both vagi then break up into branches that form the oesophageal plexus.
From the oesophageal plexus the anterior and posterior vagal trunks emerge, to enter the abdomen through the oesophageal opening in the diaphragm, and supply the stomach. The anterior vagus gives hepatic branches and the posterior vagus coeliac branches. Through these branches the vagi supply the foregut and midgut as well as their derivatives, the biliary tract and pancreas.
Accessory nerve (xi)
The spinal root of the accessory nerve is formed by fibres from cell bodies (the ‘spinal accessory nucleus’) in the anterior horn of the upper five or six segments of the cervical cord (mainly 2, 3 and 4). Unlike other anterior horn cell axons, they do not leave the cord via the anterior nerve roots, but emerge as a series of rootlets from the lateral surface of the cord behind the denticulate ligament. Joining together, they form a single nerve that ascends behind the uppermost tooth of the denticulate ligaments to join the cranial root. The latter emerges from the medulla as a series of rootlets below those of the vagus between the olive and inferior cerebellar peduncle. The spinal and cranial roots unite and the accessory nerve passes through the middle compartment of the jugular foramen, sharing the same arachnoid and dural sheath as the vagus. Outside the skull the accessory gives all its cranial root fibres to the vagus, and these are distributed by vagal branches to the striated muscle of the soft palate and larynx. The nerve then runs backwards and downwards across the internal jugular vein, as it lies in front of the transverse process of the atlas. The accessory nerve supplies sternocleidomastoid, and passes through its substance to reach the posterior triangle, which it crosses to supply trapezius.
Hypoglossal nerve (xii)
Hypoglossal nerve fibres emerge from the surface of the medulla as a vertical line of rootlets between pyramid and olive. These join into two roots that enter the hypoglossal (anterior condylar) canal in the occipital bone, where they are separated by a flange of fibrous dura mater that sometimes ossifies. They join in the canal and emerge as a single nerve.
The hypoglossal nerve passes downwards between the internal jugular vein and the internal carotid artery, until it is crossed laterally by the occipital artery and its lower sternocleidomastoid branch, as they pass backwards. The nerve then swings forwards crossing (laterally) both carotid arteries and the loop at the commencement of the lingual artery. The nerve and the lingual artery part company as they pass superficial and deep, respectively, to hyoglossus, on their way forwards to the tongue, where the nerve supplies all the muscles of the tongue except palatoglossus.
The branches of the hypoglossal nerve before it reaches the tongue are all derived from C1 nerve fibres that join the hypoglossal at its exit from the skull. A very small meningeal branch supplies dura mater in the posterior cranial fossa.
The superior root of the ansa cervicalis (formerly descendens hypoglossi) branches off as the hypoglossal nerve curves forwards from between the internal carotid artery and the internal jugular vein. It is joined by the lower root of the ansa cervicalis (formerly the descendens cervicalis) derived from C2 and C3 nerves of the cervical plexus. Together these nerves make the ansa cervicalis. The ansa usually lies on the internal jugular vein under cover of sternocleidomastoid embedded in the anterior wall of the carotid sheath. Its branches supply omohyoid, sternohyoid and sternothyroid.
The nerve to thyrohyoid comes off the hypoglossal nerve as it lies on the lingual artery, and the nerve to geniohyoid is given off in the mouth, above mylohyoid; this contains the last of the C1 fibres that travel along the hypoglossal nerve.
Part seven. Summary of cranial nerve lesions
Having relatively short courses, the peripheral parts of cranial nerves are not subject to the kind of injuries that commonly afflict peripheral nerves in the limbs. Those most commonly affected by trauma are the first, second, third, sixth and seventh, but tumours, ischaemia of nerve trunks and aneurysms of adjacent vessels are among the more usual afflictions of these and other cranial nerves.
Olfactory nerve
Head injury may tear olfactory nerve filaments passing through the cribriform plate of the ethmoid, especially if a fracture involves this part of the anterior cranial fossa. Such a fracture usually causes leakage of CSF through the nose (CSF rhinorrhoea), from tearing of the meningeal sleeves that ensheathe the olfactory nerve bundles. If all filaments on one side are torn there will be complete anosmia on that side. Test by closing one nostril (with finger pressure) and sniffing familiar substances like coffee or oranges. Most deficiencies of smell are due to affections of nasal mucosa rather than neurological disease. Smell is an essential component of taste, and complaints about lack of taste may be due to loss of smell. Olfactory hallucinations, which are usually unpleasant, arise from the uncus of the temporal lobe (the cortical centre for smell).
Optic nerve
Assessment of the visual fields tests the integrity of the visual pathways from retina to cortex, and lesions at different points along the path give rise to characteristic defects, as illustrated in Figure 7.9. The defects are conventionally described with reference to the visual fields and not to the retina. The numbers below correspond to the sites of the lesions in Figure 7.9A; clinically the most common lesions are at the chiasma (2) and in the optic radiation (4). The visual field defects are indicated in Figure 7.9B.
1.A complete lesion of the left optic nerve gives rise to complete blindness in the left eye.
2.Compression of the optic chiasma, as by a pituitary tumour, causes bitemporal hemianopia (blindness in the temporal half of both visual fields) because the nasal fibres from both retinas are interrupted. This effectively narrows the outer part of each visual field, so that the patient complains of bumping into the sides of a doorway or into people on each side.
3.A lesion of the left optic tract gives a right homonymous hemianopia, due to interruption of fibres from the same (left) sides of both retinas (hence homonymous, meaning same-sided). The field defects are therefore right-sided.
4.A lesion of the lower fibres in the left optic radiation (as from an abscess in the temporal lobe from infection spreading upwards from the middle ear) causes a right upper quadrantic homonymous hemianopia, because the lower fibres in the optic radiation are from the lower part of the retina.
5.Similar to (4), a lesion of the upper fibres in the left optic radiation (as from a parietal lobe lesion, and in practice very rare) gives a right lower quadrantic homonymous hemianopia.
6.A lesion of the anterior part of the left visual cortex (as from occlusion of the posterior cerebral artery) gives a right homonymous hemianopia similar to the optic tract lesion in (3), but there may be sparing of the macular (central) vision when the most posterior part of the visual cortex at the very tip of the occipital lobe, where macular vision is represented, is (sometimes) supplied by the middle cerebral artery.
7.Traumatic damage to the tip of the left occipital lobe, i.e. to the macular area, gives a right homonymous macular defect.
Oculomotor, trochlear and abducens nerves
Ocular nerve palsies are described on page 404. The major signs are as follows.
Oculomotor nerve: ptosis. When the lid is lifted up, the eye is looking down and out. The diplopia disappears on looking outwards. The pupil is dilated and does not react to light or on accommodation; ptosis with a large pupil thus suggests an oculomotor nerve lesion (but with a small pupil suggests Horner's syndrome; see p. 408).
Trochlear nerve : the eye cannot look down as far as it should when turned in. The head is tilted towards the opposite shoulder to compensate for extorsion.
Abducens nerve: the eye cannot look outwards.
Trigeminal nerve
Affections of the motor part of the fifth nerve, whose fibres run in the mandibular branch, are very unusual. Test for contraction of masseter.
The most common condition affecting the sensory part of the nerve is trigeminal neuralgia (tic doloureux), characterized by pain in the distribution of the maxillary and/or mandibular branches. The ophthalmic branch is rarely involved. With the maxillary nerve affected the pain is usually felt deeply in the face and nose between the mouth and orbit, and with the mandibular nerve from the mouth up to the ear and the temporal region. Compression of the trigeminal nerve adjacent to the pons by contact with a vessel (usually a branch of the basilar artery, such as the superior cerebellar or anterior inferior cerebellar artery) is considered to be a cause of trigeminal neuralgia, and patients resistant to medical therapy have been relieved of their pain surgically by the placement of a pad between the vessel and the nerve. Sclerosant injection or electrocoagulation of the trigeminal ganglion have also been utilized to abolish the pain.
The afferent side of the corneal reflex (see p. 406) depends on the ciliary branches of the nasociliary part of the ophthalmic nerve. Disappearance of the reflex is often the first sign of a lesion of the ophthalmic nerve; test by gently touching the cornea (not the conjunctiva) with cotton wool.
Facial nerve
Facial nerve paralysis is the most common of all cranial nerve lesions, and the most frequent type is Bell's palsy, a sudden onset of facial paralysis of uncertain aetiology. Paralysis of facial muscles causes asymmetry of the corner of the mouth, flattening of skin folds and inability to close the eye or wrinkle the forehead on the affected side. Test by asking the patient to show the teeth forcibly, screw up the eyes and wrinkle the forehead.
A lesion higher in the facial canal, above the origin of the chorda tympani, will add to the facial paralysis a loss of taste in the anterior part of the tongue. Test by holding the tongue out and applying sweet, sour and salt substances on cotton wool to the sides of the dorsum, asking the patient to point to the appropriate flavour written on a card. The accompanying interference with secretion from the submandibular and sublingual glands is very difficult to detect or test. A lesion higher still, above the origin of the nerve to stapedius, will give hyperacusis.
The above are all lesions of the lower motor neuron (infranuclear). A typical upper motor neuron lesion (supranuclear) paralyses the lower part of the face but not the upper (forehead and orbicularis oculi) because the upper part of the facial nerve nucleus which innervates the upper musculature is supplied by the cerebral cortex of both sides, whereas the lower part innervating the lower face only receives contralateral cortical fibres. However, emotional (as opposed to voluntary) movements of the lower facial muscles, as in smiling and laughing, are still possible with supranuclear lesions (so presumably there must be alternative pathways through the cerebrum).
Vestibulocochlear nerve
Acoustic neuromas on the extracerebral part of the eighth nerve are among the most common intracranial tumours, but other lesions of this nerve are rare. In any loss of hearing, it must be determined whether it is conduction deafness from lesions of the external or middle ear, or nerve deafness from a cochlear lesion.
The simple tests for hearing involve asking the patient to listen to whispering, the ticking of a watch, and the rubbing together of a thumb and forefinger (with the opposite ear closed by finger pressure on the tragus, and after examining the external acoustic meatus and tympanic membrane with an auroscope). Both ears are similarly tested. The basis of the two common tuning fork tests is that air conduction, which involves the amplifying effect of the ossicles across the middle ear cavity, is better than solid bone conduction. In Weber's test a vibrating tuning fork is placed on the midline of the forehead; in conduction deafness the sound is heard better in the deaf ear, and in nerve deafness it is better in the good ear. In Rinne's test the vibrating fork is held on the mastoid process until the sound is no longer heard and then quickly transferred to the external acoustic meatus; in a normal ear or with nerve deafness the sound will be heard again, but in conduction deafness further sound will not be heard. For more precise information audiometric tests are necessary.
Diseases of the internal ear or its central connections (as in Ménière's disease where there are degenerative changes in the utricle and saccule) form one group of causes of muscular incoordination or ataxia (labyrinthine ataxia; the other varieties of ataxia are cerebellar and sensory; see p. 489). The semicircular ducts of the vestibular apparatus can be tested with the head in different positions by irrigating the external acoustic meatus alternately with warm (44°C; 110°F) and cool (30°C; 86°F) water (caloric tests); this stimulates convection currents in the endolymph causing vertigo (giddiness) and nystagmus (an involuntary oscillatory movement of the eyes, which may be horizontal, vertical or rotatory). Internal ear disease gives an exaggerated response with the head in a particular position, whereas vestibular nerve damage gives an exaggerated response in any head position.
Glossopharyngeal nerve
Isolated glossopharyngeal nerve lesions are extremely rare. The last four cranial nerves may, however, be affected together (e.g. by a posterior cranial fossa tumour). Since the motor part of the glossopharyngeal nerve supplies only one small muscle, stylopharyngeus, it is impossible to test. If necessary, taste sensation on the posterior part of the tongue can be tested, with difficulty. The gag reflex (palatal elevation and pharyngeal contraction on tactile stimulation of the posterior tongue or oropharynx) tests both glossopharyngeal (afferent arc) and vagal (efferent arc) function.
Vagus nerve
Through its pharyngeal and laryngeal branches the motor component of the vagus nerve (nucleus ambiguus) supplies the pharynx, palate and larynx. Recurrent laryngeal nerve palsies are most commonly due to malignant disease (25%) and surgical damage (20%) during operations on the thyroid gland, neck, oesophagus, heart and lung. Because of its longer and partly intrathoracic course, lesions of the left nerve are more frequent than those of the right. Test the motor innervation of the vocal cords by listening to the patient's speech and cough and by inspecting the cords through a laryngoscope.
High lesions of the vagus nerve which affect the pharyngeal and superior laryngeal as well as the recurrent laryngeal branches cause difficulty in swallowing as well as vocal cord defects. The motor innervation of the soft palate can be tested by asking the patient to say a prolonged ‘Ah’ and observing the upward palatal movement; if paralysed on one side the unaffected side will rise further and pull the uvula towards the normal side.
Accessory nerve
The spinal part of the accessory nerve may be damaged in the posterior triangle, paralysing the trapezius. Test by asking the patient to shrug the shoulder; the paralysis may not be complete on account of some motor innervation occasionally reaching the muscle through cervical nerves. A lesion higher up before the nerve enters sternocleidomastoid will paralyse that muscle also; test whether the patient can turn the face to the opposite side against resistance.
Hypoglossal nerve
Paralysis of this nerve makes its own half of the tongue immobile and if of long standing the tongue will be wasted on that side. Test by asking the patient to put out the tongue; it will deviate towards the affected side due to the unopposed action of the normal half.