

29. Наследственная изменчивость. Типы мутации.
Наследственная изменчивость
Комбинативная изменчивость. Наследственную, или геноти-пическую, изменчивость подразделяют на комбинативную и мутационную.
Комбинативной называют изменчивость, в основе которой лежит образование рекомбинаций, т. е. таких комбинаций генов, которых не было у родителей.
В основе комбинативной изменчивости лежит половое размножение организмов, вследствие которого возникает огромное разнообразие генотипов. Практически неограниченными источниками генетической изменчивости служат три процесса:
Независимое расхождение гомологичных хромосом в первом мейотическом делении. Именно независимое комбинирование хромосом при мейозе является основой третьего закона Менделя. Появление зеленых гладких и желтых морщинистых семян гороха во втором поколении от скрещивания растений с желтыми гладкими и зелеными морщинистыми семенами — пример комбинативной изменчивости.
Взаимный обмен участками гомологичных хромосом, или кроссинговер (см. рис. 3.10). Он создает новые группы сцепления, т. е. служит важным источником генетической рекомбинации аллелей. Рекомбинантные хромосомы, оказавшись в зиготе, способствуют появлению признаков, нетипичных для каждого из родителей.
Случайное сочетание гамет при оплодотворении.
Эти источники комбинативной изменчивости действуют независимо и одновременно, обеспечивая при этом постоянную «перетасовку» генов, что приводит к появлению организмов с другими генотипом и фенотипом (сами гены при этом не изменяются). Однако новые комбинации генов довольно легко распадаются при передаче из поколения в поколение.
Комбинативная изменчивость является важнейшим источником всего колоссального наследственного разнообразия, характерного для живых организмов. Однако перечисленные источники изменчивости не порождают существенных для выживания стабильных изменений в генотипе, которые необходимы, согласно эволюционной теории, для возникновения новых видов. Такие изменения возникают в результате мутаций.
Мутационная изменчивость. Мутационной называется изменчивость самого генотипа. Мутации — это внезапные наследуемые изменения генетического материала, приводящие к изменению тех или иных признаков организма.
Основные положения мутационной теории разработаны Г. Де Фризом в 1901—1903 гг. и сводятся к следующему:
Мутации возникают внезапно, скачкообразно, как дискретные изменения признаков.
В отличие от ненаследственных изменений мутации представляют собой качественные изменения, которые передаются из поколения в поколение.
Мутации проявляются по-разному и могут быть как полезными, так и вредными, как доминантными, так и рецессивными.
Вероятность обнаружения мутаций зависит от числа исследованных особей.
Сходные мутации могут возникать повторно.
Мутации ненаправленны (спонтанны), т. е. мутировать может любой участок хромосомы, вызывая изменения как незначительных, так и жизненно важных признаков.
Почти любое изменение в структуре или количестве хромосом, при котором клетка сохраняет способность к самовоспроизведению, обусловливает наследственное изменение признаков организма. По характеру изменения генома, т. е. совокупности генов, заключенных в гаплоидном наборе хромосом, различают генные, хромосомные и геномные мутации.
Генные, или точковые, мутации— результат изменения нуклеотидной последовательности в молекуле ДНК в пределах одного гена. Такое изменение в гене воспроизводится при транскрипции в структуре иРНК; оно приводит к изменению последовательности аминокислот в полипептидной цепи, образующейся при трансляции на рибосомах. В результате синтезируется другой белок, что ведет к изменению соответствующего признака организма. Это наиболее распространенный вид мутаций и важнейший источник наследственной изменчивости организмов.
Существуют разные типы генных мутаций, связанных с добавлением, выпадением или перестановкой нуклеотидов в гене. Это дупликации (повторение участка гена), вставки (появление в последовательности лишней пары нуклеотидов), делеции ("выпадение одной или более пар нуклеотидов), замены нуклеотид-ных пар (AT -><- ГЦ; AT -><-; ЦГ; или AT -><- ТА), инверсии (переворот участка гена на 180°).
Эффекты генных мутаций чрезвычайно разнообразны. Большая часть из них фенотипически не проявляется, поскольку они рецессивны. Это очень важно для существования вида, так как в большинстве своем вновь возникающие мутации оказываются вредными. Однако их рецессивный характер позволяет им длительное время сохраняться у особей вида в гетерозиготном состоянии без вреда для организма и проявиться в будущем при переходе в гомозиготное состояние.
Вместе с тем известен ряд случаев, когда изменение лишь одного основания в определенном гене оказывает заметное влияние на фенотип. Одним из примеров служит такая генетическая аномалия, как серповидноклеточная анемия. Рецессивный аллель, вызывающий в гомозиготном состоянии это наследственное заболевание, выражается в замене всего одного аминокислотного остатка в (B-цепи молекулы гемоглобина (глутаминовая кислота —» —> валин). Это приводит к тому, что в крови эритроциты с таким гемоглобином деформируются (из округлых становятся серповидными) и быстро разрушаются. При этом развивается острая анемия и наблюдается снижение количества кислорода, переносимого кровью. Анемия вызывает физическую слабость, нарушения деятельности сердца и почек и может привести к ранней смерти людей, гомозиготных по мутантному аллелю.
Хромосомные мутации (перестройки, или аберрации) — это изменения в структуре хромосом, которые можно выявить и изучить под световым микроскопом.
Известны перестройки разных типов (рис. 3.13):
нехватка, или дефишенси, — потеря концевых участков хромосомы;
делеция — выпадение участка хромосомы в средней ее части;
дупликация — двух- или многократное повторение генов, локализованных в определенном участке хромосомы;
инверсия — поворот участка хромосомы на 180°, в результате чего в этом участке гены расположены в последовательности, обратной по сравнению с обычной;
транслокация — изменение положения какого-либо участка хромосомы в хромосомном наборе. К наиболее распространенному типу транслокаций относятся реципрокные, при которых происходит обмен участками между двумя негомологичными хромосомами. Участок хромосомы может изменить свое положение и без реципрокного обмена, оставаясь в той же хромосоме или включаясь в какую-то другую.
При дефишенси, делециях и дупликациях изменяется количество генетического материала. Степень фенотипического изменения зависит от того, насколько велики соответствующие участки хромосом и содержат ли они важные гены. Примеры дефишенси известны у многих организмов, включая человека. Тяжелое наследственное заболевание —синдром «кошачьего крика» (назван так по характеру звуков, издаваемых больными младенцами), обусловлен гетерозиготностью по дефишенси в 5-й хромосоме. Этот синдром сопровождается сильным нарушением роста и умственной отсталостью. Обычно дети с таким синдромом рано умирают, но некоторые доживают до зрелого возраста.
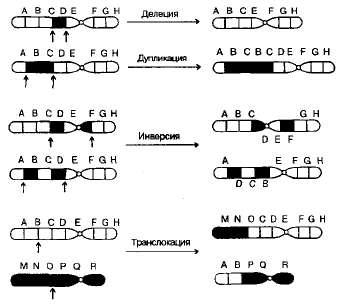
3.13. Хромосомные перестройки, изменяющие расположение генов в хромосомах.
Геномные мутации — изменение числа хромосом в геноме клеток организма. Это явление происходит в двух направлениях: в сторону увеличения числа целых гаплоидных наборов (полиплоидия) и в сторону потери или включения отдельных хромосом (анеуплоидия).
Полиплоидия — кратное увеличение гаплоидного набора хромосом. Клетки с разным числом гаплоидных наборов хромосом называются триплоидными (Зn), тетраплоидными (4n), гексанло-идными (6n), октаплоидными (8n) и т. д.
Чаще всего полиплоиды образуются при нарушении порядка расхождения хромосом к полюсам клетки при мейозе или митозе. Это может быть вызвано действием физических и химических факторов. Химические вещества типа колхицина подавляют образование митотического веретена в клетках, приступивших к делению, в результате чего удвоенные хромосомы не расходятся и клетка оказывается тетрагшоидной.
Для многих растений известны так называемые полиплоидные ряды. Они включают формы от 2 до 10n и более. Например, полиплоидный ряд из наборов в 12, 24, 36, 48, 60, 72, 96, 108 и 144 хромосомы составляют представители рода паслен (Solanum). Род пшеница (Triticum) представляет ряд, члены которого имеют 34, 28 и 42 хромосомы.
Полиплоидия приводит к изменению признаков организма и поэтому является важным источником изменчивости в эволюции и селекции, особенно у растений. Это связано с тем, что у растительных организмов весьма широко распространены гермафродитизм (самоопыление), апомиксис (партеногенез) и вегетативное размножение. Поэтому около трети видов растений, распространенных на нашей планете, — полиплоиды, а в резко континентальных условиях высокогорного Памира произрастает до 85% полиплоидов. Почти все культурные растения тоже полиплоиды, у которых, в отличие от их диких сородичей, более крупные цветки, плоды и семена, а в запасающих органах (стебель, клубни) накапливается больше питательных веществ. Полиплоиды легче приспосабливаются к неблагоприятным условиям жизни, легче переносят низкие температуры и засуху. Именно поэтому они широко распространены в северных и высокогорных районах.
В основе резкого увеличения продуктивности полиплоидных форм культурных растений лежит явление полимерии (см. § 3.3).
Анеуплоидия, или гетероплодия, — явление, при котором клетки организма содержат измененное число хромосом, не кратное гаплоидному набору. Анеуплоиды возникают тогда, когда не расходятся или теряются отдельные гомологичные хромосомы в митозе и мейозе. В результате нерасхождения хромосом при гамето-генезе могут возникать половые клетки с лишними хромосомами, и тогда при последующем слиянии с нормальными гаплоидными гаметами они образуют зиготу 2n + 1 (трисомик) по определенной хромосоме. Если в гамете оказалось меньше на одну хромосому, то последующее оплодотворение приводит к образованию зиготы 1n - 1 (моносомик) по какой-либо из хромосом. Кроме того, встречаются формы 2n - 2, или нуллисомики, так как отсутствует пара гомологичных хромосом, и 2n + х, или полисомики.
Анеуплоиды встречаются как у растений и животных, так и у человека. Анеуплоидные растения обладают низкой жизнеспособностью и плодовитостью, а у человека это явление нередко приводит к бесплодию и в этих случаях не наследуется. У детей, родившихся от матерей старше 38 лет, вероятность анеуплоидии повышена (до 2,5%). Кроме того, случаи анеуплоидии у человека вызывают хромосомные болезни.
У раздельнополых животных как в естественных, так и в искусственных условиях полиплоидия встречается крайне редко. Это обусловлено тем, что полиплоидия, вызывая изменение соотношения половых хромосом и аутосом, приводит к нарушению конъюгации гомологичных хромосом и тем самым затрудняет определение пола. В результате такие формы оказываются бесплодными и маложизнеспособными.
Спонтанные и индуцированные мутации. Спонтанными называют мутации, возникающие под влиянием неизвестных природных факторов, чаще всего как результат ошибок при воспроизведении генетического материала (ДНК или РНК). Частота спонтанного мутирования у каждого вида генетически обусловлена и поддерживается на определенном уровне.
Индуцированный мутагенез — это искусственное получение мутаций с помощью физических и химических мутагенов. Резкое увеличение частоты мутаций (в сотни раз) происходит под воздействием всех видов ионизирующих излучений (гамма- и рентгеновские лучи, протоны, нейтроны и др.), ультрафиолетового излучения, высоких и низких температур. К химическим мутагенам относятся такие вещества, как формалин, азотистый иприт, колхицин, кофеин, некоторые компоненты табака, лекарственных препаратов, пищевых консервантов и пестицидов. Биологическими мутагенами являются вирусы и токсины ряда плесневых грибов.
В настоящее время ведутся работы по созданию методов направленного воздействия различных мутагенов на конкретные гены. Такие исследования очень важны, поскольку искусственное получение мутаций нужных генов может иметь большое практическое значение для селекции растений, животных и микроорганизмов.
Закон гомологических рядов в наследственной изменчивости. Крупнейшим обобщением работ по изучению изменчивости в начале XX в. стал закон гомологических рядов в наследственной изменчивости. Он был сформулирован выдающимся русским ученым Н. И. Вавиловым в 1920 г. Сущность закона заключается в следующем: виды и роды, генетически близкие, связанные друг с другом единством происхождения, характеризуются сходными рядами наследственной изменчивости. Зная, какие формы изменчивости встречаются у одного вида, можно предвидеть нахождение аналогичных форм у родственного ему вида.
В основе закона гомологических рядов фенотипической изменчивости у родственных видов лежит представление о единстве их происхождения от одного предка в процессе естественного отбора. Поскольку общие предки имели специфический набор генов, то их потомки должны обладать примерно таким же набором.
Более того, у родственных видов, имеющих общее происхождение, возникают и сходные мутации. Это означает, что у представителей разных семейств и классов растений и животных со сходным набором генов можно встретить параллелизм — гомологические ряды мутаций по морфологическим, физиологическим и биохимическим признакам и свойствам. Так, у разных классов позвоночных встречаются сходные мутации: альбинизм и отсутствие перьев у птиц, альбинизм и бесшерстность у млекопитающих, гемофилия у многих млекопитающих и человека. У растений наследственная изменчивость отмечена по таким признакам, как пленчатое или голое зерно, остистый или безостый колос и др.
Закон гомологических рядов, отражая общую закономерность мутационного процесса и формообразования организмов, представляет широкие возможности для его практического использования в сельскохозяйственном производстве, селекции, медицине. Знание характера изменчивости нескольких родственныхх видов дает возможность поиска признака, который отсутствует у одного из них, но характерен для других. Таким путем были собраны и изучены голозерные формы злаков, односемянные сорта сахарной свеклы, не нуждающиеся в прорывке, что особенно важно при механизированной обработке почв. Медицинская наука в качестве моделей для изучения болезней человека получила возможность использовать животных с гомологическими заболеваниями: это сахарный диабет крыс; врожденная глухота мышей, собак, морских свинок; катаракта глаз мышей, крыс, собак и др.
Закон гомологических рядов позволяет также предвидеть возможность появления мутаций, еще неизвестных науке, которые могут использоваться в селекции для создания новых ценных для хозяйства форм.
Типы мутаций
Вполне вероятно, что у плодовых мушек, которых облучал Мюллер, появлялось гораздо больше мутаций, чем он смог обнаружить. По определению, мутация — это любое изменение в ДНК. Это значит, что мутации могут происходить в геноме где угодно. А поскольку большую часть генома занимает «мусорная» ДНК, ничего не кодирующая, большинство мутаций остаются незамеченными.
Мутации изменяют физические свойства организма (признаки), только если они изменяют последовательность ДНК внутри гена (рис. 7.1).

Рис. 7.1. Эти три аминокислотные последовательности показывают, как маленькие изменения могут приводить к большому эффекту. Начало одной из аминокислотных цепей в нормальном белке приведено в верхнем ряду. Ниже аминокислотная цепь ненормального варианта белка гемоглобина: валин замешен на глютаминовую кислоту в шестом положении. Эта единственная замена, приводящая к мутации кодона ГАА в кодон ГУА, является причиной серповидно-клеточной анемии, выражающейся в ряде симптомов: от слабой анемии (если у индивидуума остается нормальная копия мутировавшего гена) до смерти (если у индивидуума две мутировавшие копии гена)
Хотя Мюллер индуцировал мутации у плодовых мушек, подвергая их высоким дозам облучения, мутации случаются в организме все время. Иногда это просто ошибки нормальных процессов, происходящих в клетке, а иногда — результат воздействия окружающей среды. Такие спонтанные мутации встречаются с частотами, характерными для определенного организма, иногда называемыми спонтанным фоном.
Наиболее часто случаются точковые мутации, которые изменяют всего одну пару оснований в нормальной последовательности ДНК. Их можно получить двумя путями:
1. ДНК химически модифицируется, так что одно из оснований меняется на другое. 2. Репликация ДНК работает с ошибками, вставляя ошибочное основание в цепь при синтезе ДНК.
Какова бы ни была причина их появления, точковые мутации можно разделить на два типа:
1. Транзиции. Наиболее часто встречающийся тип мутаций. При транзиции один пиримидин замещается другим пиримидином или один пурин замещается другим пурином: например, пара Г-Ц становится парой А-Т, или наоборот.
2. Трансверзии. Более редкий тип мутаций. Пурин замещается пиримидином или наоборот: например, пара А-Т становится парой Т-А или Ц-Г.
Азотистая кислота — это мутаген, который вызывает транзиции. Она конвертирует цитозин в урацил. Цитозин обычно дает пару с гуанином, но урацил — с аденином. В результате пара Ц-Г становится парой Т-А, когда А спаривается с Т в следующей репликации. Азотистая кислота оказывает такой же эффект на аденин, превращая пару А-Т в пару Ц-Г.
Другой причиной транзиций является ошибочное спаривание оснований. Это происходит, когда по какой-то причине неправильное основание встраивается в нить ДНК, затем оно образует пару с неправильным партнером (некомплементарным основанием) вместо того, с которым должно эту пару образовать. В результате во время следующего цикла репликации пара полностью меняется.
Эффект точковых мутаций зависит от того, в каком месте последовательности оснований они образуются. Поскольку изменение одной пары оснований меняет только один кодон и, следовательно, одну аминокислоту, получающийся в результате белок может быть поврежден, но может, несмотря на повреждение, сохранить часть нормальной активности.
Гораздо сильнее, чем точковые мутации, повреждают ДНК мутации сдвига рамки. Вспомните, что генетическая последовательность оснований (секвенс) считывается как последовательность неперекрывающихся триплетов (трех оснований). Это значит, что существует три пути прочтения (рамки считывания) последовательности оснований, зависящих от точки начала прочтения. Если мутация убирает или встраивает лишнее основание, она вызывает сдвиг рамки, и вся последовательность оснований прочитывается неправильно. Это значит, что изменится вся последовательность аминокислот, а получающийся белок, с большой долей вероятности, будет полностью неработающим.
Мутации сдвига рамки вызываются акридинами, химическими веществами, которые связываются с ДНК и настолько изменяют ее структуру, что основания могут быть добавлены или убраны из ДНК во время ее репликации. Эффект таких мутаций зависит от места последовательности оснований, в котором произойдет вставка (инсерция) или выпадение (делеция) оснований, а также от их взаимного расположения в образующейся последовательности (рис. 7.2).
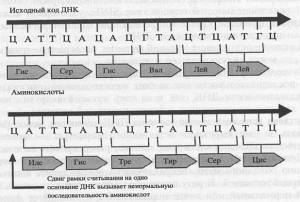
Рис. 7.2. Один из способов, которым мутация сдвига рамки может влиять на считывание последовательности оснований ДНК
Еще одним типом мутаций является встраивание (инсерция) длинных фрагментов дополнительного генетического материала в геном. Встраиваются транспозирующиеся (мобильные генетические) элементы, или транспозоны, — последовательности, которые могут перемещаться из одного места ДНК в другое. Впервые транспозоны были открыты генетиком Барбарой МакКлинток (Barbara McClintock) в 1950-е годы. Это короткие элементы ДНК, которые из одной точки генома могут перепрыгнуть в другую (поэтому их часто называют «прыгающими генами»). Иногда они прихватывают с собой расположенные рядом последовательности ДНК. Обычно транспозоны состоят из одного или нескольких генов, один из которых представляет собой ген фермента транспозазы. Этот фермент требуется транспозонам для перемещения из одного места ДНК в другое внутри клетки.
Существуют также ретротранспозоны, или ретропозоны, которые сами передвигаться не могут. Вместо этого они используют свою мРНК. Она сначала копируется в ДНК, а последняя вставляется в другую точку генома. Ретротранспозоны родственны ретровирусам.
Если транспозон встраивается в ген, кодирующая последовательность оснований нарушается, и ген в большинстве случаев выключается. Транспозоны также могут нести сигналы для окончания транскрипции или трансляции, которые эффективно блокируют выражение других генов, расположенных вслед за ними. Такой эффект называется полярной мутацией.
Ретротранспозоны типичны для геномов млекопитающих. Фактически, около 40% генома состоит из таких последовательностей. Это одна из причин, по которой геном содержит так много «мусорной» ДНК. Ретротранспозоны могут быть SINE (короткими промежуточными элементами) длиной в несколько сот пар оснований пли LINE (длинными промежуточными элементами) длиной от 3000 до 8000 пар оснований. Например, человеческий геном содержит около 300 тыс. последовательностей одного типа SINE, у которых, кажется, нет другой функции, кроме саморепликации. Данные элементы также называются «эгоистической» ДНК.
В отличие от точковых мутаций мутации, вызываемые транспозонами, не могут индуцироваться мутагенами.
Точковые мутации могут ревертировать, возвращаться к исходной последовательности как за счет восстановления оригинальной последовательности ДНК, так и за счет мутаций в других местах гена, которые компенсируют действие первичной мутации.
Вставка дополнительного элемента ДНК, очевидно, может ревертировать за счет вырезания вставленного материала — точечного исключения. Делеция части гена, однако, ревертировать не может.
Мутации могут происходить в других генах, приводя к формированию обходного пути, исправляющего повреждение, вызванное начальной мутацией. В результате образуется двойной мутант, имеющий нормальный или почти нормальный фенотип. Этот феномен называется супрессией, бывающей двух типов: внегенной и внутригенной.
Внегенная супрессорная мутация подавляет действие мутации, расположенной в другом гене, иногда за счет изменения физиологических условий, при которых белок, кодируемый супрессируемым мутантом, может функционировать вновь. Бывает, что такая мутация меняет аминокислотную последовательность мутантного белка.
Внутригенная супрессорная мутация подавляет эффект мутации в гене, где она расположена, иногда восстанавливая рамку считывания, нарушенную мутацией сдвига рамки. В некоторых случаях мутация изменяет аминокислоты в сайте, который компенсирует изменение аминокислоты, вызванное первичной мутацией. Феномен также называется реверсией во втором сайте.
Не все последовательности оснований в гене подвержены мутациям в одинаковой мере. Мутации имеют тенденцию группироваться вокруг горячих точек в последовательности гена — местах, где вероятность образования мутаций в 10 или 100 раз выше, чем ожидаемая при случайном распределении. Расположение этих горячих точек различно для разных типов мутаций и мутагенов, индуцирующих их.
В бактерии E.coli, например, горячие точки встречаются там, где расположены модифицированные основания, называемые 5-метил-цитозином. Это основание иногда подвергается таутомерному сдвигу — перестройке атома водорода. В результате Г спаривается с Т вместо Ц, а после репликации образуется пара дикого типа Г-Ц и мутантная пара А-Т (в генетике диким типом называются последовательности ДНК, которые обычно встречаются в природе).
Многие мутации не дают видимого эффекта. Они называются молчащими мутациями. Иногда мутация молчит, потому что изменения не влияют на продукцию аминокислот, а иногда — поскольку, несмотря на замену аминокислоты в белке, новая аминокислота не влияет на его функцию. Это называется нейтральной заменой.
Мутация, выключающая или изменяющая функцию гена, называется прямой мутацией. Мутация, которая реактивирует или восстанавливает функцию гена за счет реверсии начальной мутации или за счет открытия обходного пути (как при реверсии во втором сайте, описанной выше), называется обратной мутацией.
Как видите, есть много различных способов классифицировать мутации, и одна и та же мутация может относиться к различным типам. Данные табл. 7.1 могут внести ясность в характеристику мутаций.
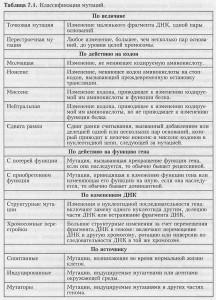
Классификация мутаций
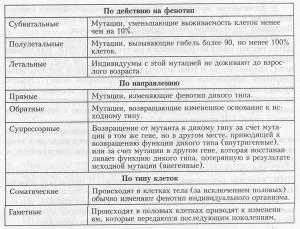
Классификация мутаций (продолжение)