

Вопрос10*Морфофункциональная организация коры больших полушарий.
Полушария большого мозга состоят из белого вещества, покрытого снаружи серым, или корой, толщина которой в различных отделах больших полушарий колеблется от 1,3 до 4,5мм. Кора представляет собой филогенетически наиболее молодой и вместе с тем сложный отдел мозга, предназначенный для обработки сенсорной информации, формирования двигательных команд и интеграции сложных форм поведения. Бурный рост неокортекса у высших позвоночных в ограниченном объеме черепа сопровождается образованием многочисленных складок, увеличивающих общую площадь коры, которая у человека составляет 2200 см2. На этом пространстве сконцентрировано 109—1010 нейронов и еще большее количество глиальных клеток, выполняющих ионорегулирующую и трофическую функции. Образующие кору нейроны по своей геометрии и функции подразделяются на несколько групп. Одну группу составляют варьирующие по размеру пирамидные клетки. Они ориентированы вертикально по отношению к поверхности коры и имеют тело треугольной формы (рис. 3.30). От тела пирамидной клетки вверх отходит длинный Т—образно ветвящийся апикальный дендрит, а вниз от основания нейрона — аксон, который либо покидает кору в составе нисходящих путей, либо направляется к другим зонам коры. Апикальные и более короткие базальные дендриты пирамидных клеток густо усеяны мелкими (до 3 мкм) выростами — шипиками, каждый из которых представляет собой область синаптического контакта.
Другая группа корковых нейронов представлена более мелкими звездчатыми клетками. Эти клетки имеют короткие сильно ветвящиеся дендриты и аксоны, формирующие внутрикорковые связи. Дендриты звездчатых клеток также могут быть снабжены шипиками, которые в процессе онтогенетического развития у человека появляются только к моменту рождения.
Наконец, третья группа корковых нейронов включает в себя веретеновидные клетки, имеющие длинный аксон, который ориентирован в горизонтальном или вертикальном направлении. В связи с тем что тела и отростки описанных выше нейронов имеют упорядоченное расположение, кора построена по экранному принципу и у млекопитающих в типичном случае гистологически состоит из шести горизонтальных слоев (рис. 3.31).
Самый наружный молекулярный слой слагается из густого сплетения нервных волокон, лежащих параллельно поверхности корковых извилин. Основную массу этих волокон составляют ветвящиеся апикальные дендриты пирамидных клеток нижележащих слоев. Сюда же в наружный слой приходят афферентные таламокортикальные волокна от неспецифических ядер таламуса, регулирующих уровень возбудимости корковых нейронов.
Второй слой — наружный зернистый — состоит из большого количества мелких звездчатых клеток, которые в вентральной части слоя дополняются малыми пирамидными клетками.
Третий слой — наружный пирамидный — формируется из пирамидных клеток средней величины. Функционально второй и третий слои коры объединяют нейроны, отростки которых обеспечивают корково—корковые ассоциативные связи.
Четвертый слой — внутренний зернистый — содержит множество звездчатых клеток (клеток—зерен), обусловливающих его гранулярную структуру. В этом слое преимущественно оканчиваются афферентные таламо—корковые волокна, идущие от специфических (проекционных) ядер таламуса.
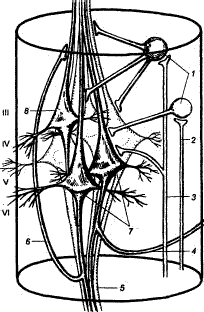
Рис. 3.30 Структурный микромодуль сенсомоторной коры
III—VI — корковые слои; 1 — интернейроны, 2 — афферентный вход, контактирующий с интернейроном, 3 — афферентный вход, образующий терминала на пирамидных нейронах, 4 — возвратная коллатераль аксона, вступающая в контакт с тормозным интернейроном, 5 — пучок аксонов, выходящий за пределы колонки, 6 — возвратная коллатераль, обеспечивающая облегчающие влияния в пределах модуля. 7 — корково—спинномозговые пирамидные клетки, 8 — корково—красноядерная пирамидная клетка.
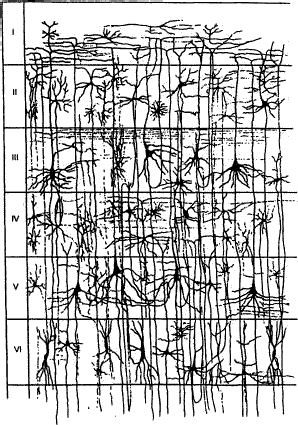
Рис. 3.31 Шестислойное строение коры большого мозга
Пятый слой — внутренний пирамидный — образован крупными пирамидными клетками. Наиболее крупные пирамидные нейроны — гигантские клетки Беца — встречаются в прецентральной извилине, занятой моторной зоной коры больших полушарий. Аксоны этих эфферентных корковых нейронов формируют корково—спинномозговой (пирамидный) и корково—бульбарный пути, участвующие в координации целенаправленных двигательных актов и позы.
И наконец, шестой слой — мультиформный, или слой веретеновидных клеток, переходящий непосредственно в белое вещество больших полушарий. Этот слой содержит тела нейронов, чьи отростки формируют корково—таламические пути.
Такой шестислойный план строения характерен для всего неокортекса (рис. 3.32). Однако выраженность отдельных слоев в различных областях коры не одинакова. Учитывая эту особенность, немецкий нейроморфолог К. Бродман по гистологическим признакам, в частности по плотности расположения и форме нейронов, разделил всю кору на 11 областей и 52 цитоархитектонических поля (рис. 3.33). Позднее были разработаны функциональные принципы классификации различных зон коры. При этом оказалось, что зоны, выделенные на основании их функциональных и нейрохимических особенностей, в известной степени соответствуют цитоархитектоническому разделению коры на поля.
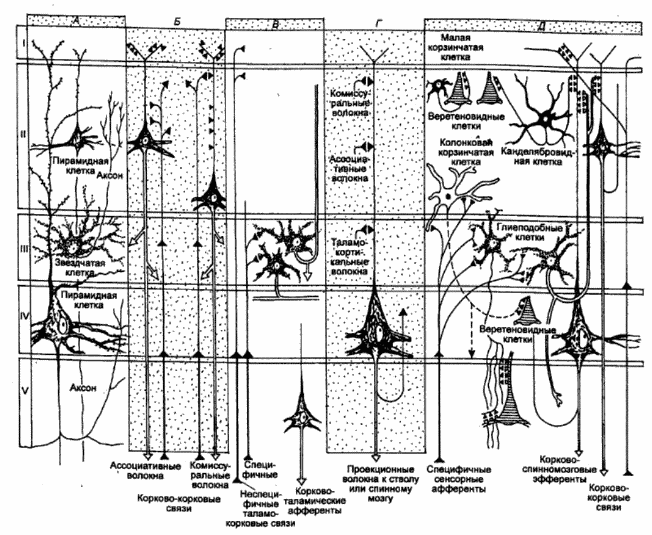
Рис. 3.32 Сильно упрощенная схема нейронов, их сетей, афферентных и эфферентных связей в различных слоях коры большого мозга
А — локализация и форма двух основных типов корковых нейронов; В — корково—корковые входы и выходы (ассоциативные а комиссуральные волокна); В — таламо—корковые (неспецифичные и специфичные) и корково—таламические связи; Г — синаптические входы пирамидной клетки, аксон которой идет к субталамическим отделам (стволу мозга и спинному мозгу); Д — обобщенная схема корковых контуров.
Так, например, при сравнении наиболее изученных сенсорных и моторных зон коры оказалось, что в первых наружный пирамидный слой выражен слабо и доминируют зернистые слои, где оканчиваются сенсорные афференты (гранулярная кора). И напротив, в моторных зонах коры зернистые слои развиты плохо (агранулярная кора), а пирамидные слои превалируют.
Таким образом, функциональная специализация накладывает определенный отпечаток на структуру сенсорных и моторных зон коры, и выделение этих областей по различным системам классификации не случайно.