

Министерство образования Пермского края
ГБОХСПО Пермский педагогический колледж физической культуры
Самостоятельная работа по дисциплине:
«Физиология с основами биохимии»
Выполнил студент группы ФК-4
Специальности
Физическая культура
Окулова Светлана
Преподаватель
Трофименкова О.Г.
Пермь 2012
Тема №1 Общая физиология возбудимых тканей
С/р: изобразить в виде схемы потенциал покоя и потенциал действия в клетке (с указанием количественной характеристики ионов Na+,Ca2+,K+,Cl-и других).
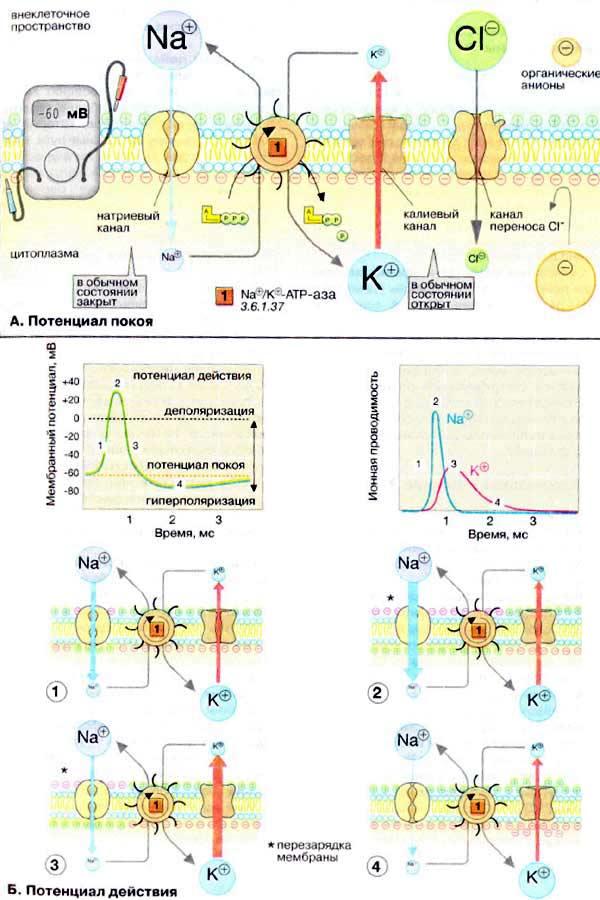
Рис.1
Деятельность Na+-канала в ходе генерации ПД можно представить следующим образом (рис. 1.). На уровне ПП А-створка закрыта, И-створка открыта (состояние готовности, точка1). При подаче стимула створки, не удерживаемые более отрицательным внутриклеточным зарядом, приходят в движение. А-створка очень быстро открывает проход внутри канала, и ионы Na+начинают входить в нейрон; натриевый ток растет (точка 2). Скоро И-створка в своем движении достигает «верхнего положения»; каналы начинают закрываться, натриевый ток ослабевает и прекращается (точка 3). При проходе уровня порога ПД совершают обратное движение створки Na+-каналов. Сначала вновь быстро срабатывает А-створка, закрывая проход канала (рис. 1, точка4), затем возвращается в исходное положение И-створка (точка 5). В результате предотвращается возможность входа дополнительной порции Na+при возврате канала в состояние готовности. Пока И-створки Na+-каналов не открылись, генерация новых ПД невозможна. Это состояние называютабсолютной рефрактерностъю.

 Рис.
2
Рис.
2
К+-каналы являются потенциал-зависимыми каналами с одной (активирующей) створкой. Относительно медленно открываясь (рис. 2, точки2—3), они обеспечивают выходящий ток калия и фазу гиперполяризации ПД. Закрытие створки происходит после «прохода» порогового уровня для запуска ПД (точки4—5). При этом скорость закрытия также невелика. В результате повышенная проницаемость мембраны для ионов К+сохраняется еще некоторое время.
В настоящее время известен не один, а несколько типов потенциал-зависимых Na+- и К+-каналов. Они различаются порогами срабатывания, а также скоростью открытия и закрытия. Кроме того, в суммарные ионные токи, возникающие при развитии ПД, значительный вклад вносят ионы Ca2+. Существуют специфические потенциал-зависимые каналы, через которые Ca2+входит в клетку; при этом он выполняет две функции: как положительно заряженный ион деполяризует мембрану и как химический агент влияет на работу других ионных каналов, ферментов, рецепторов.
Важно также то, что всякий ПД уменьшает существующую разность концентраций ионов Na+и К+внутри и снаружи клетки. Если заставить нейрон генерировать ПД с максимальной частотой, то уже через несколько тысяч импульсов за счет выхода К+и входа Na+общий заряд цитоплазмы существенно приблизится к нулю. В результате И-створки Na+-каналов не смогут открыться в конце нисходящей фазы ПД и потенциалы действия прекратятся. Теперь для восстановления работоспособности необходим отдых, и тогда Na+, К+-насосы вернут разность потенциалов к уровню ПП.
При генерации ПД возбудимость нервного волокна меняется, причем эти изменения соответствуют определенной фазе ПД.
|
№ п/п |
Фаза изменения возбудимости |
Фаза ПД |
Механизм изменения возбудимости |
Суть и значение |
|
1 |
Кратковременное повышение возбудимости |
В начале развития ПД, когда после действия раздражителя возникла некоторая деполяризация, но не достигла критического уровня (регистрируется только локальный потенциал) |
Возбудимость повышена, потому что произошла частичная деполяризация, открывается часть Na-каналов. В результате мембранный потенциал приблизился к критическому уровню деполяризации. |
|
|
2 |
Фаза абсолютной рефрактерности |
Соответствует пику ПД и продолжается в период реполяризации до достижения критического уровня деполяризации плюс-минус 10 мВ. |
В фазу деполяризации и восходящей фазы инверсии невозбудимость объясняется тем, что потенциалзависимые m-ворота Na-каналов уже открыты, остальные открываются под влиянием деполяризации, и Na быстро поступает в клетку. Поэтому дополнительное раздражение не может повлиять на движение Na. В период нисходящей фазы инверсии и реполяризации невозбудимость объясняется тем, что инактивационные h-ворота Na-каналов закрываются, в результате клеточная мембрана непроницаема для Na. В то же время в большом количестве открываются К-каналы, К быстро выходит из клетки. |
Полная невозбудимость (возбудимость = 0). Наличие абсолютной рефрактерности не позволяет отдельным ПД накладываться друг на друга и ограничивает максимальную частоту разрядов нервных клеток величиной 500—700 имп/с (реже — до 1000 имп/с: с такой частотой могут возбуждаться нейроны ретикулярной формации, толстые миелиновые нервные волокна ЦНС). |
|
3 |
Фаза относительной рефрактерности. |
Соответствует конечной части фазы реполяризации и следовой гиперполяризации до возвращения мембранного потенциала к исходному уровню после гиперполяризации. |
В фазе реполяризации повышенная проницаемости мембраны для К и избыточный выход К из клетки препятствует возможной деполяризации, поэтому нужно приложить более сильное раздражение, чтобы вызвать возбуждение. В период следовой гиперполяризации мембранный потенциал больше и, следовательно, дальше отстоит от критического уровня деполяризации, поэтому также требуется бОльший раздражитель для возбуждения. |
|
|
4 |
Фаза экзальтации |
Соответствует следовой деполяризации. |
Возбудимость повышена, т.к. вследствие частичной следовой деполяризации мембранный потенциал несколько меньше обычного и, следовательно, ближе к уровню критической деполяризации, что объясняется повышенной проницаемостью мембраны для Na. |
Период повышенной возбудимости. Возможна в тех клетках, где происходит следовая деполяризация, например, в нейронах ЦНС. Очередной ПД можно вызвать более слабым раздражением. |