

3.Витамин в6. Химическая природа, распространение, участие в обменных процессах.
Антидерматитный. Адермин, пиридоксин. Водрастворимый.
Прозводные 3-оксипиридина:
![]() пиридоксин
пиридоксин
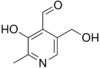 пиридоксаль
пиридоксаль
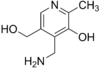 пиридоксамин
пиридоксамин
 акт.форма-
пиридоксальфосфат.
акт.форма-
пиридоксальфосфат.
Сут.потр. 2мг. Хлеб, горох, фасоль, картофель, мясо, почки, печень.
Коферментные ф.выполняют только фосфорилированные производные – ПФ. 20пиридоксалевых ферментов, катализирующих ключевые р.азотистого метаболизма в организме.
Авитаминоз:акродилия – дерматит с преим.поражением кожи лопаток, носа, ушей. Эпилептиформные припадки; дегенерация ЦНС. Гомоцистинурия, цистатионурия, нарушения обмена трп.
4.Глюкозурия и ее причины.
Обычно присутствие глюкозы в моче (глюкозурия) является результатом нарушения углеводного обмена вследствие патологических изменений в поджелудочной железе (сахарный диабет, острый панкреатит и т.д.). Реже встречается глюкозурия почечного происхождения, связанная с недостаточностью резорбции глюкозы в почечных канальцах. Как временное явление глюкозурия может возникнуть при некоторых острых инфекционных и нервных заболеваниях, после приступов эпилепсии, сотрясения мозга.
Отравления морфином, стрихнином, хлороформом, фосфором также обычно сопровождаются глюкозурией. Наконец, необходимо помнить о глюкозурии алиментарного происхождения, глюкозурии беременных и глюкозурии при нервных стрессовых состояниях (эмоциональная глюкозурия).
Билет№26
1.Обмен фенилаланина и тирозина. Использование тирозина для синтеза катехоламинов, тироксина и меланинов. Наследственные нарушения обмена фенилаланина и тирозина ( фенилкетонурия, алкаптонурия).
2.Окисление высших жирных кислот. Последовательность реакций бета-окисления. Связь окисления жирных кислот с цитратным циклом и дыхательной цепью.
Процесс окисления жирных кислот складывается из следующих основных этапов.
Активация жирных
кислот.
Свободная жирная
кислота независимо
от длины углеводородной цепи является
метаболически инертной и не может
подвергаться никаким биохимическим
превращениям, в том числе окислению,
пока не будет активирована. Активация жирной
кислоты протекает
на наружной поверхности мембраны митохондрий при
участии АТФ, коэнзима A
(HS-KoA) и ионов Mg2+. Реакция
катализируется ферментом ацил-КоА-синтетазой:![]()
В результате реакции образуется ацил-КоА, являющийся активной формой жирной кислоты.
Первая стадия дегидрирования. Ацил-КоА в митохондриях прежде всего подвергается ферментативному дегидрированию, при этом ацил-КоА теряет 2 атома водорода в α- и β-положениях, превращаясь в КоА-эфир ненасыщенной кислоты.
![]()
Стадия гидратации. Ненасыщенный ацил-КоА (еноил-КоА) при участии фермента еноил-КоА-гидратазы присоединяет молекулу воды. В результате образуется β-оксиацил-КоА (или 3-гидроксиацил-КоА):
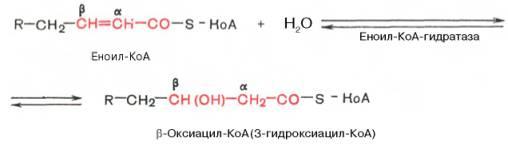
Вторая стадия дегидрирования. Образовавшийся β-оксиацил-КоА (3-гидроксиацил-КоА) затем дегидрируется. Эту реакциюкатализируют НАД+-зависимые дегидрогеназы:
![]()
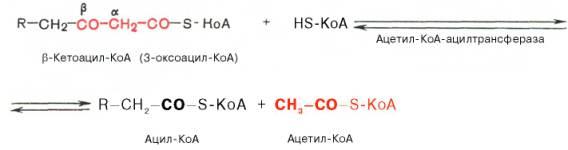
Тиолазная реакция.
представляет собой расщепление
3-оксоацил-КоА с помощью тиоловой группы
второй молекулы КоА.
В результате образуется укороченный
на два углеродных атома ацил-КоА
и двууглеродный фрагмент в виде
ацетил-КоА. Данная реакция катализируется
ацетил-КоА-ацилтрансферазой
(β-ке-тотиолазой):
Образовавшийся ацетил-КоА подвергается окислению в цикле трикар-боновых кислот, а ацил-КоА, укоротившийся на два углеродных атома, снова многократно проходит весь путь β-окисления вплоть до образования бутирил-КоА (4-углеродное соединение), который в свою очередь окисляется до 2 молекул ацетил-КоА.
Баланс энергии. При каждом цикле β-окисления образуются одна молекула ФАДН2 и одна молекула НАДН. Последние в процессеокисления в дыхательной цепи и сопряженного с ним фосфорилирования дают: ФАДН2 – 2 молекулы АТФ и НАДН – 3 молекулы АТФ, т.е. в сумме за один цикл образуется 5 молекул АТФ. При окислении пальмитиновой кислоты образуется 5 х 7 = 35 молекул АТФ. В процессе β-окисления пальмитиновой кислоты образуется 8 молекул ацетил-КоА, каждая из которых, «сгорая» в цикле трикарбоновых кислот, дает 12 молекул АТФ, а 8 молекул ацетил-КоА дадут 12 х 8 = 96 молекул АТФ.
Таким образом, всего при полном β-окислении пальмитиновой кислоты образуется 35 + 96 = 131 молекула АТФ. С учетом одноймолекулы АТФ, потраченной в самом начале на образование активной формы пальмитиновой кислоты (пальмитоил-КоА), общий энергетический выход при полном окислении одной молекулы пальмитиновой кислоты в условиях животного организма составит 131 – 1 = 130 молекул АТФ.