

56. Поток информации в клетке.
Благодаря наличию потока информации клетка на основе многовекового эволюционного опыта предков приобретает структуру, отвечающую критериям живого, поддерживает ее во времени, а также передает в ряду поколений.
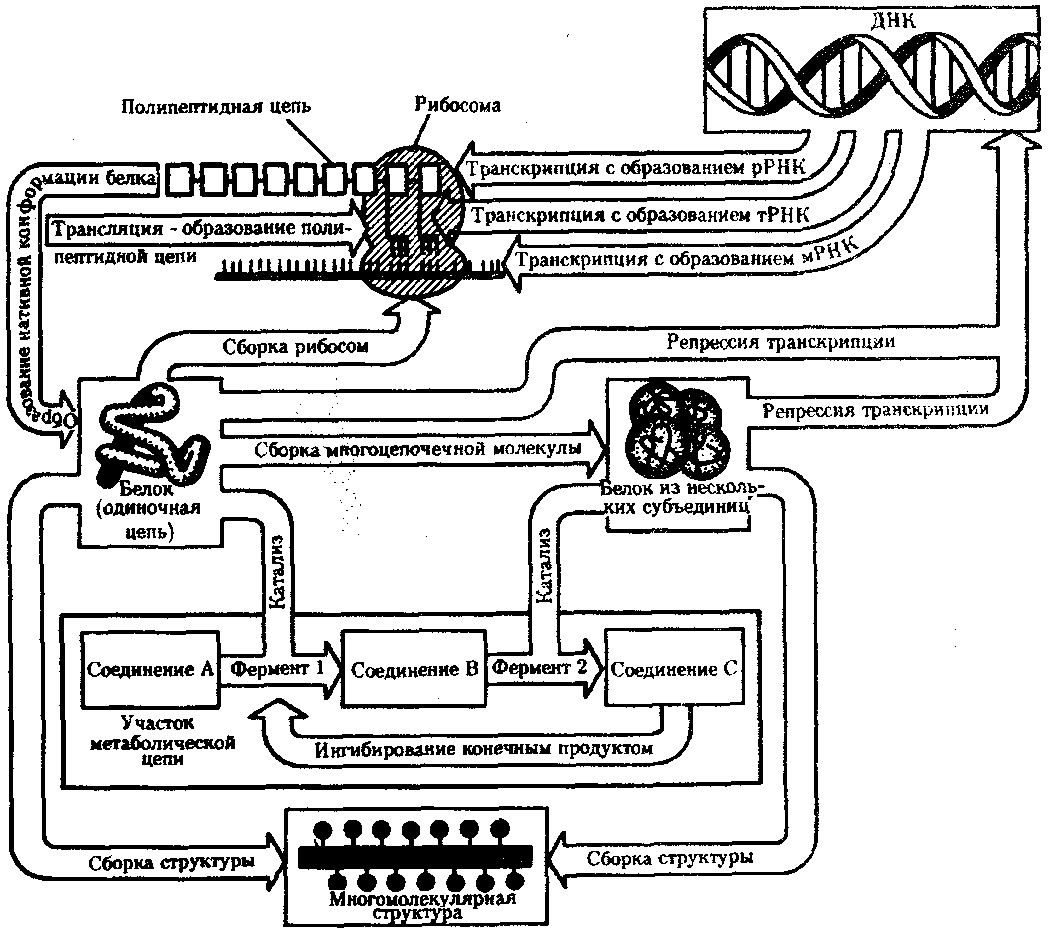
В потоке информации участвуют ядро (конкретно ДНК хромосом), макромолекулы, переносящие информацию в цитоплазму (мРНК), цитоплазматический аппарат трансляции (рибосомы и полисомы, тРНК, ферменты активации аминокислот). На завершающем этапе этого потока полипептиды, синтезированные на полисомах, приобретают третичную и четвертичную структуры и используются в качестве катализаторов или структурных белков. Кроме основного по объему заключенной информации ядерного генома в эукариотических клетках функционируют также геномы митохондрий, а в зеленых растениях — и хлоропластов.
66. Матричный синтез как специфическое свойство живого.
Матричный cинтез — реакции полимеризации и поликонденсации, при которых строение образующегося полимера и (или) кинетика процесса определяются другими макромолекулами — матрицами, находящимися в непосредственном контакте с молекулами одного или нескольких мономеров и растущими цепями.
Пример матричного синтеза в живой природе — биосинтез нуклеиновых кислот и белков, в котором роль матрицы играют ДНК и РНК, а состав и порядок чередования остатков мономеров в синтезируемом полимере однозначно определяются строением матрицы.
Термин «матричный синтез» обычно используют при описании синтеза нуклеиновых кислот и белков. При рассмотрении способов получения других полимеров используют иные термины — матричные полиреакции, полимеризация, поликонденсация. Такой матричный синтез реализуется при условии химического и пространственного соответствия (комплементарности) мономеров и растущей цепи, с одной стороны, и матрицы — с другой; при этом элементарные акты осуществляются между мономерами и растущими макромолекулами (а также олигомерами — при матричной поликонденсации), связанными с матрицей.
Обычно мономеры и олигомеры обратимо связываются с матрицей достаточно слабыми межмолекулярными взаимодействиями — водородными, электростатическими. Дочерние цепи в некоторых случаях практически необратимо ассоциируют с матрицей («узнают» матрицу) только после того, как достигнут некоторой определенной длины, зависящей от энергии взаимодействия между звеньями матрицы и дочерней цепи. «Узнавание» матрицы растущей цепью — необходимая стадия матричного синтеза; дочерние цепи практически всегда содержат фрагмент или фрагменты, образовавшиеся по «обычному» механизму, то есть без влияния матрицы. Скорость матричный синтез может быть выше, ниже или равна скорости процесса в отсутствие матрицы (кинетический матричный эффект).
Структурный матричный эффект проявляется в способности матрицы влиять на длину и химическое строение дочерних цепей (в том числе их пространственную структуру), а если в матричном синтезе участвуют два или более мономера — то также на состав сополимера и способ чередования звеньев.
76. Регуляция экспрессии генов у прокариот. Индукция синтеза катаболических ферментов(Lac-оперон).
Регуляция экспрессии генов у прокариот.
У прокариот известно несколько механизмов регуляции экспрессии генов: изменение дозы генов, регуляция на уровне транскрипции, посттрансляционная модификация полипептидов и т.д. Наиболее распространена регуляция на уровне транскрипции, которая по сравнению с другими способами регуляции очень экономна.
В 1961 французские ученые Франсуа Жакоб, Жак Люсьен Моно и Андре Мишель Львов предложили гипотезу оперона, объясняющую регуляцию синтеза белков на уровне транскрипции у прокариот.
Оперон – это совокупность тесно сцепленных структурных генов прокариот вместе с участком-оператором, регулирующим их транскрипцию. Структурные гены несут информацию о структуре белков, необходимых для выполнения одной функции. К оперону также относятся регуляторные участки: промотор и терминатор, которые участвуют в управлении транскрипцией.
Промотор – это участок ДНК, определяющий начало транскрипции. С промотором связывается РНК-полимераза, которая затем продвигается вдоль ДНК, синтезируя молекулу РНК.
Терминатор – это участок ДНК, обозначающий окончание транскрипции. РНК-полимераза, дойдя до терминатора, заканчивает транскрипцию и покидает ДНК.
Оператор – это участок ДНК, с которым может специфически связываться белок-репрессор. Информацию о белке-репрессоре содержит ген-регулятор, который не входит в состав оперона. Связавшись с оператором, репрессор останавливает транскрипцию, т.к. препятствует продвижению РНК-полимеразы вдоль ДНК.
Когда оперон активен, РНК-полимераза транскрибирует все структурные гены на одну молекулу иРНК. Информационная РНК, содержащая несколько генов, называется полицистронной.
Индукция синтеза катаболических ферментов(Lac-оперон).
Лактозный оперон включает в себя 3 структурных гена, которые несут информацию о ферментах, расщепляющих лактозу. В отсутствие лактозы ферменты, ее расщепляющие, не синтезируются. Синтез этих ферментов останавливается на стадии транскрипции. Транскрипцию блокирует белок-репрессор, который связывается с оператором.
Индуктором, который включает синтез ферментов, является лактоза, которая способна связываться с репрессором. Когда в клетке появляется лактоза, она связывается с белком-репрессором и переводит его в неактивную форму. Неактивный репрессор теряет способность связываться с ДНК и уходит с оператора. РНК-полимераза получает возможность транскрибировать структурные гены. В результате транскрипции образуется иРНК, содержащая 3 структурных гена. На каждом гене происходит трансляция, в результате которой синтезируются ферменты, расщепляющие лактозу.
По мере расщепления лактозы ее концентрация в клетке падает, белок-репрессор освобождается от лактозы, переходит в активную форму, вновь приобретает способность связываться с ДНК, садится на оператор и останавливает трансляцию.