

Биология ДНК
.docx1. ДНК была открыта Иоганном Фридрихом Мишером в 1868 году. Из остатков клеток, содержащихся в гное, он выделил вещество, в состав которого входят азот и фосфор. Вначале новое вещество получило название нуклеин, а позже, когда Мишер определил, что это вещество обладает кислотными свойствами, вещество получило название нуклеиновая кислота. Биологическая функция новооткрытого вещества была неясна, и долгое время ДНК считалась запасником фосфора в организме. Более того, даже в начале XX века многие биологи считали, что ДНК не имеет никакого отношения к передаче информации, поскольку строение молекулы, по их мнению, было слишком однообразным и не могло содержать закодированную информацию.
Постепенно было доказано, что именно ДНК, а не белки, как считалось раньше, является носителем генетической информации. Одно из первых решающих доказательств принесли эксперименты О. Эвери, Колина Мак-Леода и Маклин Мак-Карти (1944 г.) по трансформации бактерий. Им удалось показать, что за так называемую трансформацию (приобретение болезнетворных свойств безвредной культурой в результате добавления в неё мёртвых болезнетворных бактерий) отвечают выделенная из пневмококков ДНК.
Пневмококки в норме образуют гладкие (то есть крупные, с ровной поверхностью) колонии и имеют полисахаридную капсулу, компоненты которой и запускают образование антител.
В ходе эксперимента пневмококки, образующие гладкие колонии, были убиты нагреванием, и из них был извлечён компонент, растворимый в водно-солевом растворе. Белки были осаждены хлороформом, а полисахаридные капсулы, обусловливающие антигенные свойства бактерий, гидролизованы специфичным ферментом. Для подтверждения полного гидролиза капсул, была проведена процедура иммунопреципитации специфическими антителами. После разделения в спирте из полученной активной фракции были выделены волокнистые тяжи[2].
Химический анализ показал, что соотношение углерода, водорода, азота и фосфора в полученном осадке соответствует соотношению этих же элементов в молекуле ДНК. Для подтверждения того, что действующим началом трансформации является именно ДНК, но не РНК, белки или другие компоненты клетки, Эвери с сотрудниками обработали смесь трипсином, химотрипсином, рибонуклеазой, но эта обработка никак не влияла на трансформирующие свойства. Лишь обработка ДНКазой приводила к разрушению трансформирующего начала. Таким образом было установлено, что действующим началом бактериальной трансформации является дезоксирибонуклеиновая кислота (ДНК).
Эксперимент американских учёных Алфреда Херши и Марты Чейз (эксперимент Херши — Чейз, 1952 г.) с помеченными радиоактивными изотопами белками и ДНК-бактериофагов показали, что в заражённую клетку передаётся только нуклеиновая кислота фага, а новое поколение фага содержит такие же белки и нуклеиновую кислоту, как исходный фаг.
2. Мономер ДНК — нуклеотид (дезоксирибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания нуклеиновых кислот относятся к классам пиримидинов и пуринов. Пиримидиновые основания ДНК (имеют в составе своей молекулы одно кольцо) — тимин, цитозин. Пуриновые основания (имеют два кольца) — аденин и гуанин.
3. Молекула ДНК образована двумя полинуклеотидными цепями, спирально закрученными друг около друга и вместе вокруг воображаемой оси, т. е. представляет собой двойную спираль (исключение — некоторые ДНК-содержащие вирусы имеют одноцепочечную ДНК) . Диаметр двойной спирали ДНК — 2 нм, расстояние между соседними нуклеотидами — 0,34 нм, на один оборот спирали приходится 10 пар нуклеотидов. Длина молекулы может достигать нескольких сантиметров. Молекулярный вес — десятки и сотни миллионов. Суммарная длина ДНК ядра клетки человека — около 2 м. В эукариотических клетках ДНК образует комплексы с белками и имеет специфическую пространственную конформацию.
4. Молекула ДНК обладает несколькими свойствами: 1) во-первых, она удваивается (процесс называется репликация и происходит во время S периода интерфазы. При этом копии ДНК поровну распределяются в дочерние клетки в период митоза) - передача наследственной информации; 2) во-вторых, она реализует свою генетическую информацию. Этот процесс называется транскрипцией, в ходе транскрипции с ДНК снимается копия в виде РНК, а уже РНК непосредственно участвует в синтезе белков - реализация наследственной информации; 3) в случае повреждения ДНК может восстановить себя, что называется репарацией, данный процесс поддерживает стабильность генетической информации - стабильное хранение наследственной информации; Итого 3 свойства: репликация, транскрипция, репарация.
5. Репликация ДНК — ключевое событие в ходе деления клетки. Принципиально, чтобы к моменту деления ДНК была реплицирована полностью и при этом только один раз. Это обеспечивается определёнными механизмами регуляции репликации ДНК. Репликация проходит в три этапа:
-
инициация репликации
-
элонгация
-
терминация репликации.
Регуляция репликации осуществляется в основном на этапе инициации. Это достаточно легко осуществимо, потому что репликация может начинаться не с любого участка ДНК, а со строго определённого, называемого сайтом инициации репликации. В геноме таких сайтов может быть как всего один, так и много. С понятием сайта инициации репликации тесно связано понятие репликон. Репликон — это участок ДНК, который содержит сайт инициации репликации и реплицируется после начала синтеза ДНК с этого сайта. Геномы бактерий, как правило, представляют собой один репликон, это значит, что репликация всего генома является следствием всего одного акта инициации репликации. Геномы эукариот (а также их отдельные хромосомы) состоят из большого числа самостоятельных репликонов, это значительно сокращает суммарное время репликации отдельной хромосомы. Молекулярные механизмы, которые контролируют количество актов инициации репликации в каждом сайте за один цикл деления клетки, называются контролем копийности. В бактериальных клетках помимо хромосомной ДНК часто содержатся плазмиды, которые представляют собой отдельные репликоны. У плазмид существуют свои механизмы контроля копийности: они могут обеспечивать синтез как всего одной копии плазмиды за клеточный цикл, так и тысяч копий[1].
Репликация начинается в сайте инициации репликации с расплетания двойной спирали ДНК, при этом формируется репликационная вилка — место непосредственной репликации ДНК. В каждом сайте может формироваться одна или две репликационные вилки в зависимости от того, является ли репликация одно- или двунаправленной. Более распространена двунаправленная репликация. Через некоторое время после начала репликации в электронный микроскоп можно наблюдать репликационный глазок — участок хромосомы, где ДНК уже реплицирована, окружённый более протяжёнными участками нереплицированной ДНК.
В репликационной вилке ДНК копирует крупный белковый комплекс (реплисома), ключевым ферментом которого является ДНК-полимераза. Репликационная вилка движется со скоростью порядка 100 000 пар нуклеотидов в минуту у прокариот и 500—5000 — у эукариот.
Молекулярный механизм репликации
Ферменты (хеликаза, топоизомераза) и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований и активностью ДНК-полимеразы, способной распознать и исправить ошибку. Репликация у эукариот осуществляется несколькими разными ДНК-полимеразами. ДНК-полимераза I действует на запаздывающей цепи для удаления РНК-праймеров и дорепликации очищенных мест ДНК. ДНК полимераза III — основной фермент репликации ДНК, осуществляющий синтез ведущей цепи ДНКи фрагментов Оказаки при синтезе запаздывающей цепи. Далее происходит закручивание синтезированных молекул по принципу суперспирализации и дальнейшей компактизации ДНК. Синтез энергозатратный.
Цепи молекулы ДНК расходятся, образуют репликационную вилку, и каждая из них становится матрицей, на которой синтезируется новая комплементарная цепь. В результате образуются две новые двуспиральные молекулы ДНК, идентичные родительской молекуле.
Характеристики процесса репликации
-
матричный — последовательность синтезируемой цепи ДНК однозначно определяется последовательностью материнской цепи в соответствии с принципом комплементарности;
-
полуконсервативный — одна цепь молекулы ДНК, образовавшейся в результате репликации, является вновь синтезированной, а вторая — материнской;
-
идёт в направлении от 5’-конца новой молекулы к 3’-концу;
-
полунепрерывный — одна из цепей ДНК синтезируется непрерывно, а вторая — в виде набора отдельных коротких фрагментов (фрагментов Оказаки);
-
начинается с определённых участков ДНК, которые называются сайтами инициации репликации (англ. origin).
6. Генети́ческий код — свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов.
Свойства
-
Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон).
-
Непрерывность — между триплетами нет знаков препинания, то есть информация считывается непрерывно.
-
Неперекрываемость — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов (не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки).
-
Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте (однако, кодон UGA у Euplotes crassus кодирует две аминокислоты — цистеин и селеноцистеин).
-
Вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов.
-
Универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусов до человека (на этом основаны методыгенной инженерии; есть ряд исключений, показанный в таблице раздела «Вариации стандартного генетического кода» ниже).
-
Помехоустойчивость — мутации замен нуклеотидов, не приводящие к смене класса кодируемой аминокислоты, называют консервативными; мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными.
8.
Особенности организации генома эукариот
У эукариотических организмов механизм регуляции транскрипции гораздо более сложен. В результате клонирования и секвенирования генов эукариот обнаружены специфические последовательности, принимающие участие в транскрипции и трансляции.
Для эукариотической клетки характерно:
1. Наличие интронов и экзонов в молекуле ДНК.
2. Созревание и-РНК - вырезание интронов и сшивка экзонов.
3. Наличие регуляторных элементов, регулирующих транскрипцию, таких как: а) промоторы - 3 вида, на каждый из которых садится специфическая полимераза. РНК-полимераза I реплицирует рибосомные гены, РНК-полимераза II - структурные гены белков, РНК-полимераза III - гены, кодирующие небольшие РНК. Промоторы РНК-полимеразы I и РНК-полимеразы II находятся перед участком инициации транскрипции, промотор РНК-полимеразы III - в рамках структурного гена; б) модуляторы - последовательности ДНК, усиливающие уровень транскрипции; в) (энхансеры) усилители - последовательности, усиливающие уровень транскрипции и действующие независимо от своего положения относительно кодирующей части гена и состояния начальной точки синтеза РНК; г) терминаторы - специфические последовательности, прекращающие и трансляцию, и транскрипцию.
Эти последовательности по своей первичной структуре и расположению относительно инициирующего кодона отличаются от прокариотических, и бактериальная РНК-полимераза их не "узнает". Таким образом, для экспрессии эукариотических генов в клетках прокариот нужно, чтобы гены находились под контролем прокариотических регуляторных элементов. Это обстоятельство необходимо учитывать при конструировании векторов для экспрессии.
Структурные гены, обеспечивающие жизнедеятельность эукариотической клетки, обычно транскрибируется в большинстве активно функционирующих клеток. В то же время, специфические гены, уникальные тех или иных тканей или органов, транскрибируются и транслируются только в определенных клетках. Например, гены, кодирующие α- и β-субъединицы гемоглобина взрослого человека, экспрессируются исключительно в клетках - предшественниках эритроцитов. Число разных мРНК, специфичных для разных клеток, варьирует от единиц до десятков.
Способность клеток включать (активировать) или выключать (ингибировать) структурные гены крайне важна для поддержания клеточной специфичности и экономного расходования энергетических ресурсов. Отсюда и многообразие факторов транскрипции, имеющих белковую природу. Многие из них связываются непосредственно с нуклеотидной последовательностью длиной менее 10 п.н., называемой по-разному: боксом, модулем, элементом инициации, регуляторным элементом. В отличие от прокариот у эукариот опероны в большинстве своем отсутствуют, т. е. каждый эукариотический структурный ген имеет свой собственный набор регуляторных элементов. Существенную роль в регуляции транскрипции у эукариот, помимо опосредованной взаимодействием между ДНК и белками, играют также белок-белковые взаимодействия. Несмотря на индивидуальность набора регуляторных элементов у структурных генов эукариот, каждый из них имеет промоторный участок (ТАТА-бокс, или бокс Хогнесса) из восьми нуклеотидов, включающий последовательность TATA; последовательность ССААТ (САТ-бокс); участок из повторяющихся динуклеотидов GC (GC-бокс). Эти элементы находятся на расстоянии 25, 75 и 90 п.н. от сайта инициации соответственно:
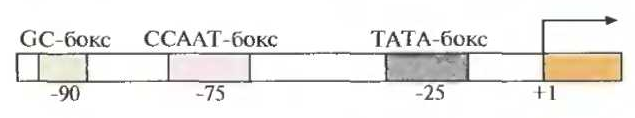 Регуляторные
элементы структурных генов эукариот.
Отрицательное значение показывает, что
эти элементы находятся в молекуле ДНК
слева от сайта инициации транскрипции,
обозначенного +1. Стрелка — направление
транскрипции (по Глик Б. , Пастернак,
Дж., 2002)
Регуляторные
элементы структурных генов эукариот.
Отрицательное значение показывает, что
эти элементы находятся в молекуле ДНК
слева от сайта инициации транскрипции,
обозначенного +1. Стрелка — направление
транскрипции (по Глик Б. , Пастернак,
Дж., 2002)
Транскрипция структурного гена эукариот начинается со связывания с ТАТА-боксом фактора транскрипции, который представляет собой комплекс по крайней мере из 14 белков. Затем с ним и участками ДНК, примыкающими к ТАТА-боксу, связываются другие факторы транскрипции, и, наконец, со всем этим транскрипционным комплексом связывается РНК-полимераза II. Затем при участии дополнительных факторов происходит инициация транскрипции в точке +1 . Если последовательность TATA отсутствует или существенно изменена, то транскрипция структурного гена становится невозможной.
9. В конце 60-х годов работами американских ученых Р. Бриттена, Э. Дэвидсона и других была открыта фундаментальная особенность молекулярной структуры генома эукариот – нуклеотидные последовательности разной степени повторяемости. Это открытие было сделано с помощью молекулярно-биологического метода изучения кинетики ренатурации денатурированной ДНК. Различают следующие фракции в геноме эукариот.
1.Уникальные, т.е. последовательности, представленные в одном экземпляре или немногими копиями. Как правило, это цистроны – структурные гены, кодирующие белки.
2.Низкочастотные повторы – последовательности, повторяющиеся десятки раз.
3.Промежуточные, или среднечастотные, повторы – последовательности, повторяющиеся сотни и тысячи раз. К ним относятся гены рРНК (у человека 200 на гаплоидный набор, у мыши – 100, у кошки – 1000, у рыб и цветковых растений – тысячи), тРНК, гены рибосомных белков и белков-гистонов.
4. Высокочастотные повторы, число которых достигает 10 миллионов (на геном). Это короткие (~ 10 пн) не кодирующие последовательности, которые входят в состав прицентромерногогетерохроматина.
У эукариот объем наследственного материала значительно больше. В отличие от прокариот в эукариотических клетках одновременно активно транскрибируется от 1 до 10% ДНК. Состав транскрибируемых последовательностей и их количество зависят от типа клетки и стадии онтогенеза. Значительная часть нуклеотидных последовательностей у эукариот не транскрибируется вообще — молчащая ДНК.
Большой объем наследственного материала эукариот объясняется существованием в нем помимо уникальных также умеренно и высоко повторяющихся последовательностей. Эти высоко повторяющиеся последовательности ДНК располагаются в основном в гетерохроматине, окружающем центромерные участки. Они не транскрибируются. Характеризуя наследственный материал прокариотической клетки в целом, необходимо отметить, что он заключен не только в нуклеоиде, но также присутствует в цитоплазме в виде небольших кольцевых фрагментов ДНК —плазмид.
Плазмиды — это широко распространенные в живых клетках внехромосомные генетические элементы, способные существовать и размножаться в клетке автономно от геномной ДНК. Описаны плазмиды, которые реплицируются не автономно, а только в составе геномной ДНК, в которую они включаются в определенных участках. В этом случае их называют эписомами.
В прокариотических (бактериальных) клетках обнаружены плазмиды, которые несут наследственный материал, определяющий такие свойства, как способность бактерий к конъюгации, а также их устойчивость к некоторым лекарственным веществам.
В эукариотических клетках внехромосомная ДНК представлена генетическим аппаратом органелл — митохондрий и пластид, а также нуклеотидными последовательностями, не являющимися жизненно необходимыми для клетки (вирусоподобными частицами). Наследственный материал органелл находится в их матриксе в виде нескольких копий кольцевых молекул ДНК, не связанных с гистонами. В митохондриях, например содержится от 2 до 10 копий мтДНК.
Внехромосомная ДНК составляет лишь небольшую часть наследственного материала эукариотической клетки.