

Лихенометрия
.docЛихенометрия
Лишайники
Лихенометрический метод датирования
Индексы возраста
Функции роста лишайников-индикаторов
Построение кривых роста с учетом старения локальных лишайниковых популяций
Лишайники
|
Лишайники представляют собой симбиотические организмы, состоящие из 3 компонентов - микобионта (гриба), сине-зеленой водоросли и азотофиксирующей бактерии. Они широко распространены в современной биосфере во всех физико-географических зонах. Даже на поверхности ледникового щита в центральной Гренландии небольшой метеорит, обнаруженный спустя несколько десятилетий после падения, оказался колонизованным накипными лишайниками. Лишайники имеют несколько экологических типов - эпилитные, т. е. растущие только на камнях, эпифитные - на растениях, некоторые виды специализируются только на костях животных, другие на металле, третьи на минералах и породах определенного состава и т.д. Кроме того, лишайники по морфологии слоевища или таллома разделяются на накипные (пленочные), корковые, чешуйчатые, листоватые и кустистые. Лишайниковые сообщества представляют собой уникальную альтернативную модель биосферы и являются наиболее замкнутыми и самодостаточными среди других наземных экосистем. |
Космополитный арктроальпийский накипный лишайник Rhizocarpon Alpicola |

|
Лишайники, по сравнению с другими макроорганизмами имеют наиболее широкую экологическую амплитуду и большое видовое разнообразие. Симбиотический организм лишайника является как продуцентом, так и редуцентом органического вещества и кислорода. Лишайники колонизируют абиотическую среду, преобразуют и меняют ее свойства и вместе с микроорганизмами, увеличивая разнообразие среды, создают новые ниши для более сложных экосистем. Появление данных симбиотических организмов, а возможно и целой их наземной биосферы, вероятно, является результатом одного из “экспериментов выхода жизни” на сушу несколько миллиардов лет назад. Причиной отсутствия ископаемых свидетельств древних лишайников является их низкая биомасса, к тому же наиболее устойчивые к разрушению части лишайников - споры несут лишь информацию о простейших одноклеточных организмах, следы присутствия которых известны в архейских отложениях. Скорости роста разных лишайников существенно различаются. Относительно быстрорастущими являются некоторые тропические кустистые виды, ежегодный прирост которых достигает более 10 см. Намного медленнее растут тонкие накипные циркумарктические виды, которые наиболее часто используются в лихенометрии, например роды Rhizocarpon, Aspicilia, Leucanora, и др. Скорости роста особей одних и тех же видов зависят от конкретных физико-географических условий и возраста самих особей. |
|
Лихенометрический метод датирования Исходными предпосылками для использования лишайников в качестве индексов возраста, а точнее времени экспонирования конкретной морфоскульптурной поверхности являются: 1) большая продолжительность жизни отдельных особей некоторых видов, достигающая в суровых условиях Арктики и Субарктики более 5-7 тыс. лет; 2) “стремление” талломов к равномерному радиальному росту; 3) сглаживание аномально высоких годовых приростов, обусловленных мелко периодическими климатическими флуктуациями, внутри большого периода жизни особей. Современное состояние метода, его преимущества и недостатки отражены в ряде публикаций Р.Е. Бешеля (R.Beschel), П. Ворсли (P.Worsley), Дж. Иннеса (J.Innes), Дж. Андрюса и П. Веббера (J. Andrews, P.Webber), Дж. Бенедикта (J.Benedict), П. Беркланда (P.Birkeland), А.А.Никонова и Т.Ю.Шебалиной, О.С.Савоскула, О.Н. Соломиной, К.И. Турманиной, А.А.Галанина и ряда других исследователей. |
Особь циркумполярного эпилитного вида Rhizocarpon Alpicola - в центре; слившиеся особи Rhizocarpon секции Rhizocarpon - по краям, Parmelia sp. - округлые черные |
|
Индексы возраста В преобладающем объеме лихенометрических исследований в качестве индекса возраста используется диаметр наиболее крупной особи лишайника-индикатора. Поэтому существенную долю в ошибки оценок времени экспонирования вносит дисперсия скорости роста отдельных слоевищ популяции, зависящая от множества экологических, популяционно-ценотических, микроклиматических и других факторов. Внесение поправок на их влияние в алгоритм расчета практически невозможно. Некоторые исследователи в качестве индекса возраста предлагают использовать выборочную статистику, полученную путем осреднения некоторого количества наиболее крупных особей. Однако этот вариант также малопригоден вследствие весьма асимметричного распределения лишайниковых популяций на большинстве датируемых поверхностей. Ненормальные частотные распределения диаметров талломов лишайников-индикаторов ограничивают доверительность к лихенометрическим оценкам возраста, полученных путем использования осредненных диаметров, поскольку в этом случае не учитываются ошибки. |
|
|
|
|



|
|
|
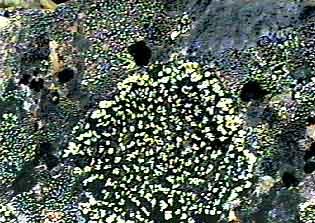
Частотные распределения лишайников индикаторов на морфоскульптурных поверхностях разного генезиса и современной динамики . По оси X - интервал группирования; по оси Y - диаметры талломов в мм |

|
Поэтому в качестве возрастных индексов нами предложены параметры логарифмического тренда наложенного на убывающий вариационный ряд выборки диаметров талломов поверхности. Эти ряды с высокой достоверностью (R^2>0,8) описываются уравнениями вида Y=aLnX+b, где Y - порядковый номер таллома в ряду, X - ожидаемый его диаметр, а, b - коэффициенты. Значение коэффициента b предсказывает диаметр теоретического таллома, который мог быть не обнаружен во время проведения измерений.
Функции роста лишайников-индикаторов Основное количество лихенометрических оценок проводится на основе линейной зависимости между диаметром самого большого слоевища и минимальным возрастом датируемой поверхности. Однако еще Р. Бешелем установлено, что ежегодные приросты лишайников по мере старения замедляются и, вероятно, останавливаются вообще. Иначе лишайники могли бы достигать невероятно крупных размеров. Р.Е. Бешелем были выделены фаза быстрого нелинейного роста и, следующая за ней, линейная фаза. Однако, вряд ли существует какое-то физиологическое свойство накипных лишайников, обусловливающее внезапный переход от нелинейного к линейному росту. |

|
Наиболее оптимальным способом оценки функции роста лишайников-индикаторов является трендовый анализ эмпирической кривой роста, построенной на основе репрезентативной выборки с реперных лихенометрических площадок, датированных независимыми абсолютными методами. Построение
таких уравнений на практике оказалось
весьма затруднительным из-за отсутствия
достаточного количества реперных
поверхностей в одном
ландшафтно-географическом районе.
Причем некоторые исследователи
продолжают использовать линейные
зависимости. Например, одно из уравнений
О.Н.Соломиной, построенное для
лишайников из рода Rhizocarpon в одном из
район t = 24,8 + 13,5dRh, где t - минимальный возраст поверхности, dRh - диаметр наиболее крупной особи Rhizocarpon sp. Для лихенометрических оценок Дж. Метьюс [Matthews, 1974; 1975; 1977] вывел уравнение, использующее логарифмическую функцию log(t+c)=a+bx, где t - возраст поверхности, x - диаметр максимальной особи |
Теоретическая кривая роста лишайника Rhizocarpon geographicum для районов северной Норвегии, построенная по зависимости log(y+c)=a+bx Дж. Метьюса [Matthews, 1977] |
 ов
Тянь-Шаня имеет вид:
ов
Тянь-Шаня имеет вид:
|
лишайника-индикатора, а, b и с - коэффициенты, зависящие от особенностей используемого вида и физико-климатических условий района датирования. Очевидно, что для нахождения этих коэффициентов необходимо решить систему из трех уравнений. Для территории Норвегии этим исследователем коэффициенты а, b и с были оценены в 1,8143, 0,0062 и 50 соответственно. В предлагаемой автором зависимости не возраст, а логарифм возраста линейно связан с диаметром максимального лишайника. Выполненный нами анализ данной зависимости при разных значениях x показал, что она отражает представления Р.Е. Бешеля о фазе быстрого нелинейного роста и, следующей за ней линейной фазе. Однако, по данной функции в интервале возрастов более 100 лет, она принимает форму, близкую к линейной. Что противоречит условию об предельных размерах роста слоевищ накипных лишайников. А.А.Никоновым с соавторами для построения вероятностной модели роста локальных популяций лишайника индикатора использовался известный закон замедления роста численности живых организмов - Т.Р. Мальтуса. Профессионально математически постороенная авторами модель весьма сложна для практического использования, к тому же в ее основе приняты некоторые априорные допущения, противоречащие законам конкуренции и популяционно-ценотических отношений в локальных сообществах накипных лишайников [Галанин, 1997; 1999]. Тем не менее, среди известных лихенометрических методик она является наиболее удовлетворительной, позволяя рассчитать ошибки возрастных оценок. В качестве замедляющей функции нами был принята убывающая экспонента и допущение, что замедление прироста максимального теоретического диаметра локальной выборки происходит по экспоненциальному закону. Пусть в первый год после заселения какой-либо “свежей” каменной поверхности прирост используемой статистики (максимального теоретического диаметра) лишайников-индикаторов составил ао мм/год. Тогда средний выборочный прирост аi в год i с учетом экспоненциального замедления будет: ai= aо * e^ (-i / f), где f - некий параметр замедления. Максимальный теоретический диаметр dt на поверхности возраста t лет будет равен сумме годовых приростов: t dt =a0* e^ (-i / f) , i= 1 а после интегрирования t dt a0*e^ (-i / f)(di) = a0f*(e^ (-1 / f) - e^ (-i / f)) . 1 Величина e^ (-1 / f) уже при f >>10 приближается к 1, следовательно, для приближенных расчетов можно использовать формулу: d a0f * (1 - e^ (-i / f))(1) . На поверхности с небольшим временем экспонирования замедление роста слоевищ существенно не проявится, поэтому: dt a0t (2). Можно показать, что при большом возрасте каменной поверхности (t ) величина e^ - t/f 0, следовательно, в этих случаях будет верна формула dt a0 f (3), где dt - средний выборочный диаметр на поверхности большого (условно бесконечного) времени экспонирования. Предлагаемая методика позволяет построить приближенную кривую роста для района, в котором имеется 1 или более независимо датированных поверхностей колонизованных накипными лишайниками. Начальный годовой прирост приближенно вычисляется по формуле 2, используя результаты измерений талломов на поверхности известного возраста, например горной выработке, карьере, с использованием дендрохронологических данных и др. Параметр замедления роста рассчитывается, используя приближение 3, по результатам изменений диаметров слоевищ на поверхности древнего условно бесконечного возраста. Такие поверхности хорошо определяются по степени выветрелости обломков, характеру склоновых процессов, лишайниковым сукцессиям и другим геоморфологическим признакам. Параметры роста в разных районах неодинаковы и зависят от их физико-географических условий, определяющих продолжительность вегетационного периода лишайников. Для субарктической тундры побережья Анадырского лимана параметры ао и f для среднего значения ста максимальных особей соответственно составляют: секция Rhizocarpon - 0,14±0,01 мм/год и 526±95 лет, род Aspicilia - 0,15±0,01 мм/год и 733±115 лет. В лесотундровой зоне центральной части Корякского нагорья для секции Rhizocarpon - 0,18±0,03 мм/год и 452±88 лет, а начальный годовой прирост листоватого лишайника Parmelia sp. - 0,47±0,08 мм/год. В Примагаданье (зона северной тайги) параметры роста лишайников рода Rhizocarpon секции Rhizocarpon составляют 0,23±0,03 мм/год и 419±86 лет. Таким образом, скорости роста исследуемых лишайников в разных физико-географических районах существенно различаются. Наиболее низкие годовые приросты и, соответственно, более медленный рост и развитие популяций накипных лишайников характерно для побережья Анадырского лимана, что не противоречит условиям субарктического климата. Параметры роста накипных лишайников в Северной Корякии и Примагаданье близки по значениям, что, по-видимому, можно объяснить близкими климатическими характеристиками этих районов. |
|
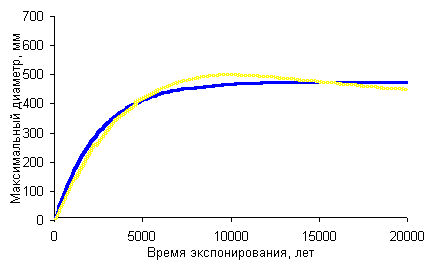
Формула 3 отражает то, что спустя определенный период времени экспонируемая поверхность и обитающие на ней лишайники “стареют”. Наступает фаза равновесия, когда прирост наиболее крупных диаметров слоевищ лишайника-индикатора останавливается. На рисунке справа видно, что значения максимального диаметра ограничено горизонтальной асимптотой (синяя кривая). Для практического применения пригодна левая нелинейная часть кривой со значениями возраста менее 7000-5000 лет. Ошибки вычисления исходных параметров и возраста по данной методике связаны с величиной достоверности аппроксимации вариационных рядов используемых выборок, возраста реперной поверхности реперной поверхности, а также приближенным характером используемой экспоненциальной функции. Метод расчета ошибок лихенометрических параметров роста и возраста при использовании в качестве возрастного индекса выборочного среднего нами приведен ранее [Галанин, 1997]. Средняя ошибка |
Синяя линия - кривая роста Rhizocarpon geographicum для хребта Брукс (Аляска), построенная нами путем итераций по формуле 1 и данным П.Калкина и Дж.Эллиса [Calkin, Ellis, 1980]; желтая линия - график ее полиноминального тренда |
|
|
|
датирования по этой методике составляет около 20%.
|
Кривая роста Rhizocarpon sp. для гипсометрического интервала 200-400 м Северо-охотского побережья. Аппроксимационные тренды различных функций и разной степенью приближения. Для датирования в районе наиболее приемлемо уравнение полинома 4-й степени |
В виду сложностей с поиском датированных реперных поверхностей для построения лихенометрических кривых на начальном этапе исследований для приближенных оценок мы построили кривые для циркумполярно распространенного лишайника Rhizocarpon sp. для некоторых районов Северо-Востока, используя средне выборочные значения диаметров особей на поверхностях, где лишайники-индикаторы достигают своих предельных размеров [Галанин, 1996; 1997]. В 1999 г. были найдены два реперных объекта в черте г. Магадана на гипсометрических отметках 200-400 м датированных историческими данными и по С14, что позволило путем решения системы из 2-х уравнений 1 более точно оценить параметры роста Rhizocarpon sp. и построить лихенометрическую кривую для этого района (рисунок слева). Рассчитанные параметры составили: начальный прирост а0 - 0,29 мм/год; замедление f - 1183 года. |

![]()
|
Построение кривых роста с учетом старения локальных лишайниковых популяций Наблюдения за популяциями эпилитных лишайников показали, что наиболее крупные особи обитают не на самых древних плакорных поверхностях, а на субстрате более молодого возраста. Наиболее крупные лишайники, обитающие на поверхностях, время экспонирования которых превышает этот возрастной оптимум, имеют меньшие диаметры [Галанин, 1996, 1999]. Это объясняется постепенными сукцессионными сменами, замещением эдификаторов и доминантов в локальном сообществе по мере выветривания и разрушения поверхности морфоскульптуры, изменения морфологических и химических свойств субстрата. Поэтому истинная кривая роста наиболее часто используемых эпилитных видов, например, из рода Rhizocarpon, должна описываться полиномиальной зависимостью и иметь математический экстремум, отвечающий субстратно-возрастному оптимуму. |
|
Кривая роста Rhizocarpon sp. для нивально-гляциальных районов Корякского хребта и Чукотского п-ова [Галанин, 1999]. 1 - реперные датировки, 2 - положения неогляциальных образований ледника “Цирк” в бассейне р. Находка (Корякский хребет) |
Эти положения были нами подтверждены при исследовании позднеплейстоценовых и голоценовых ледниковых валунов, время экспонирования которых было определено космоизотопным методом [Глушкова, Гуалтиери, 1999; Галанин, 1999; Gualtiery, Glushkova, 2000]. На основании нескольких датированных альтернативными методами реперных поверхностей нами была построена лихенометрическая кривая Rhizocarpon sp. для нивально-гляциальных ландшафтов Корякского хребта, учитывающая влияние старения популяции на уменьшение используемого индекса возраста: b=-0,000003t^2+0,056t (4), где b - значение максимального теоретического диаметра (коэффициент логарифмической регрессии), t - время экспонирования поверхности [Галанин, 1999]. Уравнение 4 пригодно для проведения приближенного лихенометрического датирования гляциальной и перигляциальной каменной морфоскульптуры на Чукотке. |

|
Ошибка датирования по данному уравнению составляет 5-15% и зависит от времени экспонирования датируемой поверхности и точности оценки теоретического максимального диаметра. Выведенная лихенометрическая кривая отражает различные этапы популяционной динамики лишайников Rhizocarpon секции Rhizocarpon на локальной каменной поверхности с момента ее колонизации, когда особи данного рода являются доминантами в сообществе (левая и средняя части кривой), до стадии сукцессионной смены и вытеснения Rhizocarpon sp. другими доминантами из родов Aspicilia и Haematomma (правая часть кривой). Данная закономерность популяционно-ценотической динамики установлена нами на основе сравнения совместных частотных распределений выборок из этих трех родов, являющихся эдификаторами в сообществах накипных лишайников в перигляциальных ландшафтах Северо-Востока [Галанин, 1997; 1999]. Поэтому при проведении лихенометрического датирования по данной кривой необходимо учитывать, что “стадия доминирования” Rhizocarpon секции Rhizocarpon может длиться от 3 до 6 тыс. лет, в зависимости от степени устойчивости экспонируемой породы к выветриванию и денудации. В этом временном интервале особи из родов Aspicilia и Haematomma имеют намного меньшие морфометрические показатели, а толщина выветрелого слоя экспонируемой породы под их слоевищами не превышает 0,5-1 см. После завершения стадии “доминирования” и развития выветрелой корки породы более 1-2 см происходит постепенная сукцессионная смена, и доминантами в локальных сообществах накипных лишайников становятся Aspicilia sp. и Haematomma sp. Особи из рода Rhizocarpon в силу морфологических и физиологических особенностей слоевищ не могут скреплять и удерживать шелушащиеся и выпадающие обломки экспонируемой породы и постепенно вытесняются лишайниками с более толстыми слоевищами, гифы которых проникают в глубь выветрелой корки, скрепляют ее, приостанавливая тем самым процесс шелушения. Поэтому рыхлые слоевища Haematomma ventosum, достигающие толщины более 1 см, как правило, обильно насыщены песком, дресвой и даже мелким щебнем. При сильном выветривании, когда толщина выветрелой корки становится более 1-2 см, морфометрические показатели (максимальный и средний диаметры) локальной популяции Rhizocarpon sp. уменьшаются в 2 раза и более по сравнению с предшествующим периодом доминирования. На таких поверхностях сукцессионные смены затухают и практически останавливаются, наступает состояние динамичного равновесия (климакса). Время, необходимое для достижения данного равновесия, определяется как индивидуальными особенностями выветриваемой горной породы (главным образом денудационная устойчивость), так и системой местных физико-климатических и экологических условий. По результатам наблюдений локальных популяций указанных выше родов на верхнеплейстоценовых и голоценовых моренах в горах Чукотки нами установлено, что в подавляющем большинстве случаев для достижения климаксного состояния в сообществе эпилитных лишайников требуется 7-10 тыс. лет экспонирования каменной поверхности. Таким образом, при проведении лихенометрического датирования с использованием выведенного нами уравнения кривой для Rhizocarpon секции Rhizocarpon необходимо учитывать качественные лихенометрические характеристики датируемой поверхности, приведенные в таблице 1. Данной таблицей можно пользоваться и для “грубого” экспрессного отнесения каменных поверхностей перигляциальных ландшафтов по обитающим на них лишайникам и их размерам к нижнему, среднему или современному интервалам голоцена. |