

The Art of Genes How Organisms Make Themselves
.pdfDo not despise my opinion, when I remind you that it should not be hard for you to stop sometimes and look into the stains of walls, or ashes of a fire, or clouds, or mud or like places, in which, if you consider them well, you may find really marvellous ideas.
When Leonardo looks at a series of stains and sees a landscape with a battle in progress, he is clearly making a rather individual interpretation. Someone else looking at the same set of stains might interpret them in an entirely different way. But it is this very ambiguity in the stains, their ability to be interpreted in different ways, that can lead the observer to make new discoveries. If he was advising someone on how to paint a landscape, why not look at a real landscape rather than a stained wall? Unlike a real landscape, the greater ambiguities in the wall allow for a free ranging and creative mind to see novel forms that might not be arrived at by the more obvious routes. In a limited sense the forms are already there on the wall to begin with: once the shape of a face is pointed out, it is there for all to see. Nevertheless, the act of staring at the wall and coming up with particular shapes dearly adds something, because one response is selected out of an enormous possible number. Interpretation is therefore not a neutral event, but a highly selective process.
The same principle, put to a different purpose, underlies the use of ink blots in a psychological test, described by H. Rorschach in the 1920s. In this test, a person is shown a series of ink blots and asked to interpret them in whatever way he or she chooses. Not all ink blots will do for the test: they are purposely chosen to be suggestive but ambiguous, in order to stimulate various types of interpretation. Perhaps on being shown the blot in Fig. 3.7, someone might see a butterfly or a bat, or something else.
Now the responses to particular blots are dearly not random. If they were random— bearing no relation whatever to the material being shown— we would not call them interpretations but simply random utterances. The tricky part is trying to explain what leads someone to select one particular interpretation. If a person is shown the ink blot in Fig. 3.7 and says it looks like a bat, we can be reasonably confident that he has previously seen a bat or at least a picture of a bat. But most people also know what butterflies look like, so why might someone say 'bat' instead of 'butterfly'? In some cases we might be able to come up with a reasonable answer, such as that he saw a vampire movie the previous night and bats were therefore on his mind. But in most cases it would be difficult to decide. (The Rorschach test avoids this problem because it does not depend on trying to account for particular responses. The test is based on a compilation of interpretations given by 'normal' subjects, and those given by people with various psychological problems. Diagnosis proceeds by seeing whether a subject's profile of responses falls within or outside the 'normal' range, as defined by previous compilations.)
 Fig.3.7 Ink blot.
Fig.3.7 Ink blot.
The factors that lead to one interpretation over another can be so subtle and complex as to defy solution. The best general comment we could make is that a subjects response has to do with his or her state of mind. This in turn depends on how the subject's brain has developed and responded to previous experiences, going back to childhood and beyond. In other words, the response is historically informed: it depends in some way on the biological and environmental past of the individual. But this is so complicated, involving so many interactions, contingencies and chance events, that finding an explanation for one response as compared to another may effectively be impossible. This is true whether we are talking about ink blots, stains on a wall, clouds or any other highly ambiguous shape.
These examples serve to illustrate three key features of interpretations: (1) In each case we can identify a common frame of reference that might be shown to anyone, some stains on a wall or an ink blot. (2) Individuals respond to this common information in a selective way, giving one response amongst many. (3) The selections are not random but are historically informed, depending on the individual's biological and cultural past, usually in a very complex way.
We can now return to the question of how a gene can be said to interpret something. Like the other examples of interpretations, the various bristle patterns conferred by the scute gene depend on a common frame of reference, the pattern of hidden colours. This is the same in each fly, irrespective of what the final bristle pattern looks like. Based on this common underlying patchwork, each version of the scute gene then ensures that only one specific pattern of bristles is produced; it selects one outcome amongst many. The process therefore resembles interpretations in two key respects. I now need to explore possible parallels with the third feature of interpretations: the historical process that leads the scute gene to respond in one way rather than another.
The evolution of pattern
Why do organisms display particular patterns, such as a specific arrangement of bristles, rather than others? As with tracing the origin of mental interpretations, this is a complex historical problem. To illustrate this, it will be helpful to switch from bristles to a different type of pattern which is much better understood in terms of its evolution: the colour patterns of butterfly wings.

In 1848, the naturalists Henry Walter Bates and Alfred Russel Wallace left England on a joint expedition to explore the Amazon. Wallace returned after four years, but Bates stayed on for a further seven. During his long stay, Bates managed to accumulate an enormous number of biological specimens, including about 14000 species of insects. Bates was particularly struck by a curious feature of the butterflies: there was a surprisingly dose resemblance in colour patterns between some distantly related species, suggesting to him that one group of butterfly species might be mimicking the other. Because some of the species were thought to be distasteful to predators, he proposed that other species might mimic them to fool predators into thinking that they were distasteful too. This is illustrated in Fig. 3.8, taken from his paper, where the bottom row shows two brightly coloured species that birds quickly learn to avoid eating as they contain noxious compounds and are therefore unpalatable. The butterflies in the top row look very similar superficially to those below them, but they are all palatable varieties of a distantly related species that birds are able to eat without ill effect. However, because they mimic the unpalatable species, these varieties will also be avoided by birds, based on their previous learning experiences. The palatable species is effectively hitching a ride on the learned response of birds to the other species (this only works effectively if the mimetic species is not too common relative to the unpalatable species: otherwise the birds would mostly encounter palatable mimics and would not learn avoidance). This phenomenon is called Batesian mimicry after its first proponent.
Fig. 3.8 Unpalatable species of butterfly (bottom row) and their palatable mimics (top row). The species shown are: Leptalis amphione (top left), Leptalis orise (top right), Mechanitis polyrnnia (bottom left) and Methona psidii (bottom right) (from Bates 1862 ).
But why has this particular strategy evolved? Why does the palatable species go to all the trouble of mimicking the other, rather than just evolving an unpleasant taste of its own? To answer this question we have to look at why some butterflies taste unpleasant. In many cases the nasty taste comes from their having eaten noxious plant compounds during their earlier life as caterpillars. Based on this, we can come up with a plausible scenario for how unpalatability might have evolved. It starts with a particular species or group of plants that produce a noxious chemical to protect themselves from being eaten by animals. Being well protected, the plants thrive. Eventually, an insect species overcomes the plants' defence, by sequestering the poisonous chemicals out of harm's way, say in a particular gland. The insect is now free to gorge itself on a

normally harmful group of plants without competition and it therefore spreads very successfully. As an added bonus, the insect is now harmful to any other animal that tries to eat it because of the stored chemical. This is one way, amongst other possibilities, in which some nasty-tasting butterflies may have evolved.
We are therefore faced with two possible evolutionary outcomes: in one case the butterfly acquires a nasty taste, in the other case it mimics the appearance of an unpalatable species. Which of these is more likely to occur?
The evolution of mimicry depends on several conditions. To begin with, there have to be heritable variations, mutations, leading to a chance resemblance between the palatable and unpalatable species that tends to fool a bird every so often. Although the initial resemblance might be very poor, natural selection acting on a series of such variations over many generations might eventually give a closer and closer match. Whether this series of events occurs depends on how different the species are to begin with and how mutations happen to affect wing patterns. There are also further issues that have to be borne in mind. For instance, in some butterfly species, male colour is thought to be used in signalling to other rival males. Thus if a mutation that changed colour pattern also reduc ed the ability of a male to compete, any advantage it might gain from mimicry could be outweighed by the disadvantage to mating. In other words, a change in pattern that is advantageous from one point of view will only be selected for if it does not jeopardise other important roles. Whether mimicry evolves is therefore contingent on a range of circumstances. There are many cases in which one or more of these will not be fulfilled, and for this reason it is perhaps not too surprising that mimicry has not evolved in many butterfly species, even though they are palatable.
What about the evolution of a nasty taste? In the scenario I gave, a caterpillar has to overcome a plant's defence by sequestering a noxious chemical. As with the evolution of mimicry, this would involve a series of mutations arising by chance and being acted upon by natural selection. Whether this is at all feasible in this case would depend on factors like the digestive system of the insect, the availability of a suitable site for sequestration, the insect's normal feeding pattern and how mutations happen to affect all of these processes. Only with the appropriate conditions might such a mechanism evolve.
Whether or not evolution will lead to a butterfly species being a mimic or acquiring a nasty taste (or perhaps neither of these, or even both), therefore depends on a constellation of factors concerning the species and its environment. The colour pattern we see displayed by a butterfly today is clearly not random, it is based on the past. But because evolution proceeds by modifications of what Went before, the particular route it has taken will have been affected by numerous biological interactions, contingencies and chance events.

Fig. 3.9 Silk brocade with Arabic inscription, thirteenth or fourteenth century. Victoria and Albert Museum, London
Many of the same principles apply to man-made patterns. A good illustration of this comes from Archibald Christie's book, Pattern Design. By the early Middle Ages, the art of silk-weaving had achieved great perfection in many Islamic countries. It was common practice at these times for the Muslim silk weavers to insert expressions of good-will in their fabrics, following an ancient tradition. The piece of silk shown in Fig. 3.9 includes an Arabic inscription of this type reading 'GLORY, VICTORY, AND PROSPERITY'.
As the art of silk-weaving spread westwards, other countries started to imitate the Muslim fabrics, including the various inscriptions. Being incomprehensible in medieval Europe, the written characters eventually degenerated through successive copying into meaningless scribbles (Fig. 3.10). Various stages in this degeneration can be followed in the silk fabrics themselves, as well as in representations of them in paintings or carvings. In the carving of a veil, shown in the right part of Fig. 3.10, the artist seems to have been dissatisfied with the confused pattern and tried to bring it nearer to convention by making it look more like western script. Some early writers on Gothic art spent fruitless hours trying to decipher the meaning of these strange characters, not realising that they were debased versions of Arabic.
Fig. 3.10
Debased Arabic inscriptions. On the left is a woven silk fabric: Sicily, late thirteenth century. On the rig ht is part of a border carved on a veil worn by a sculptured figure: France, fifteenth century.
In some cases, the instinctive desire for order and harmony led to further modifications of the letters into symmetrical patterns, known as 'mock Arabic' devices, as shown in the upper three silk fabrics in Fig. 3.11. Other developments resulted in a complete loss of any resemblance to script, and led to a distinctive type of pattern, as shown in the bottom part of Fig. 3.11.
Fig. 3.11
Debased Arabic inscriptions in various stages of formalisation. The top three patterns are from Italian silks of the fourteenth century. The bottom pattern comes from an initial letter in an illuminated manusc ript Bible at Winchester Cathedral, England, late twelfth century.
Now if you were simply presented with some of the later versions of debased Arabic, without any knowledge of their origins, they might strike you as little more than attractive patterns. They may seem to have been devised purely on the basis of their immediate appeal. We only get a
deeper understanding by tracing their idiosyncratic past, their history of migration from a culture that happened to use one form of script to a different culture that copied the patterns without comprehension of their original meaning. Through successive copying, errors crept in and alterations were introduced, eventually leading to distinctive patterns. Although the later designs look quite different, they are still imbued with the influence of the original script.
Genes and interpretations
These examples of the evolution of butterfly patterns and human designs serve to show how patterns can change through a series of modifications, each step depending on what went before. The same can be said for fly bristles. The typical pattern of bristles displayed by a fruit fly depends on a complicated evolutionary trail, to which a whole range of different circumstances will have contributed. It is in this sense that we can say that the normal response of the scute gene to the hidden patchwork of colours is historically informed. The scute gene reacts in one way amongst many other possibilities because of a complex evolutionary past. We shall deal with precisely how this sort of response can evolve in a later chapter.
We can summarise by saying that the scute gene affects the bristle pattern by making a particular interpretation. Like other examples of interpretation, this involves making a selective and informed response to a common frame of reference, the pattern of hidden colours in the fly. As we shall see, the ability to interpret hidden colours is not just a feature of scute, it is a property of almost all the genes in an organism. Once appreciated, this will provide an essential key to understanding how the language of genes operates during development. But to get any further, we will first need to take a closer look at the underlying patterns of hidden colours.

Chapter 4 A case of mistaken identity
At three in the morning on the third of September, 1786, Johann Wolfgang Goethe jumped into a coach, assumed a false name, and set off for Italy. Goethe had just turned 37. In his youth, he had achieved great success with the publication of a tragic novel, The Sorrows of Young Werther. The book was so popular that a cult industry rapidly grew around it. There were Werther plays, operas and songs; even pieces of porcelain were made showing Werther scenes. In spite of his outstanding literary success, Goethe chose at the age of 26 to serve for a period in the court of Weimar, at the invitation of the Duke. At various times during the next eleven years he assumed responsibilities for the mines, the War Department, and the Finances of the Duchy. However, life in Weimar eventually proved too restrictive and by the time he was 37 Goethe felt impelled to escape incognito to a new environment.
Goethe travelled around Italy for about twenty months. During this time he developed various scientific theories concerning the weather, geology and botany. It may come as a surprise that so famous a poet should have concerned himself with science. Goethe, though, had far-ranging interests in nature. His scientific work was particularly important to him, and he dedicated much of his time to it. The aspect that most concerns us here is an important botanical idea he had during his Italian journey.
A unifying theme
To understand Goethe's idea and how he came to it we need to go back a few years to a discovery he made during his period at Weimar at the age of 34. Goethe had been struck by fundamental similarities in the structures of different organisms and became convinced that they were all formed in a common way. One of the most obvious illustrations of this was the similar arrangements of bones in the skeletons of many different animals. For instance, the human thigh bone, or femur, had an easily identified counterpart in a dog, bull, lion or any other mammal. However, although such a one-to-one correspondence could be established for most bones in the body, there were some apparent exceptions. For example, monkeys had a bone in the middle of their face, called the intermaxilla, which appeared to be lacking in humans (this bone is also known as the premaxillary). This was often taken to be an important distinguishing mark that separated man from ape. But Goethe's belief in a fundamental unity between organisms encouraged him to look much more closely at the human skull. Eventually he discovered that the inter-maxillary bone was also present in man but it had been overlooked because it was tucked away in the upper jaw and was closely joined with other bones (Fig. 4.1). His conviction in the commonality of forms had led him to discover something that others had missed. He was able to show that rather than being a distinguishing mark, the intermaxillary bone was actually a connecting link that unified man with other animals.
Fig. 4.1
Intermaxillary bone of human and monkey
One piece of evidence that Goethe used to support his identification of the bone came from abnormalities. He noted that in individuals born with a cleft palate, the cleft almost always ran along the join between the proposed inter-maxillary region and the surrounding bones, pointing to the intermaxillary bone as being a separate entity. He was using a rare congenital defect, the deft palate, as a way of more dearly revealing what was normally going on. It was a type of argument he was to employ again in support of his botanical theories.
During his time at Weimar, Goethe also developed a profound interest in botany, helped by teachers from the nearby Academy at Jena. The local forests, gardens and estates provided an extensive flora on which he could practice and apply his botanical knowledge. But it was only when he went to Italy that a unifying idea about plants started to crystallise, as he explained in an autobiographical essay later in life:
everything that has been round about us from youth, with which we are nevertheless only superficially acquainted, always seems ordinary and trivial to us, so familiar, so commonplace that we hardly give it a second thought. On the other hand, we find that new subjects, in their striking diversity, stimulate our intellects and make us realise that we are capable of pure enthusiasm; they point to something higher, something which we might be privileged to attain. This is the real advantage of travel and each individual benefits in proportion to his nature and way of doing things. The well-known becomes new, and, linked with new phenomena, it stimulates attention, reflection and judgement.
Exposed to a new flora during his Italian journey, Goethe was stimulated to think more deeply about plants. As with his work on skulls, he was searching for a fundamental unity that lay behind the surface of things. He came to realise that there was a single underlying theme to plants, epitomised by the leaf. It seemed to him that the same theme occurred again and again throughout the life of every plant:
While walking in the Public Gardens of Palermo, it came to me in a flash that in the organ of the plant which we are accustomed to call the leaf lies the true Proteus* (*Proteus is a sea god of Greek and Roman mythology fabled to assume various shapes.)who can hide or reveal himself in all vegetal forms. From first to last, the plant is nothing but leaf, which is so inseparable from the future germ that one cannot think of one without the other. Anyone who has had the experience of being confronted by an idea, pregnant with possibilities, whether he thought of it for himself or caught it from others, will know that it creates a tumult and enthusiasm in the mind, which makes one intuitively anticipate its further developments and the conclusions towards which it points.
Knowing this, he will understand that my vision had become an obsessive passion with which I was to be occupied, if not exclusively perhaps, still for the rest of my life.
On returning to Germany, Goethe wrote up his idea in an essay, The Metamorphosis of Plants, published in 1790. He began by describing the typical life of a plant. After germination of the seed, a tiny shoot bearing one or two small leaves emerges from the ground. As the seedling grows, foliage leaves are successively produced, spaced out around the axis of the stem. At this stage all there is to the plant is stem and leaves (Goethe was not concerned with roots in his account).
Eventually, however, the plant starts to form flowers. The question was how flowers might be related to the rest of the plant. Goethe proposed that the different parts of a flower were fundamentally equivalent to foliage leaves; it was just that instead of being spaced out along a stem, the parts of a flower were all clustered together.
Take a flower and look at its component parts. You will find that there are several types of
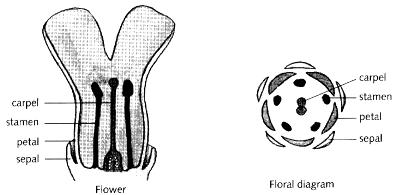
organs, clustered around each other in concentric rings or whorls (Fig. 4.2). By whorl, I mean a region or zone of the flower that normally includes organs of one type (this is not quite the same as a botanist's definition but it will be more useful for our purposes). Many flowers have four whorls of organs. The outermost whorl comprises the sepals, usually small green leaf-like structures that protect the flower when it is in bud. Within these is a whorl of petals, usually the most obvious and attractive parts of a flower. Next come the stamens, the male sex organs that bear pollen. Finally, in the centre are the carpels, the female organs that when pollinated will grow to form fruits containing seeds. This concentric arrangement is shown diagrammatically in the right part of Fig. 4.2.
Fig. 4.2 Section through a typical flower (left) showing organs arranged in concentric whorls of sepals, petals, stamens and carpels. The overall arrangement is also shown in diagrammatic form (right).
Goethe proposed that the floral organs, as well as all the foliage leaves, were simply different manifestations of a common underlying theme. This theme could be realised in different ways during plant growth, first as foliage leaves, then as the organs of a flower: sepals, petals, stamens and carpels. It seemed as though an underlying organ was simply passing through a series of different forms. He called this process of change metamorphosis, by analogy with the changes many insects experience (though unlike insect metamorphosis where the whole organism undergoes a change, Goethe's version is more abstract and refers only to parts of the organism expressing a change, the various leaf-like organs). Accordingly, above ground level, a plant was made solely of stems and a series of different types of organs based on a common theme.
In support of his claim, Goethe emphasised the many similarities between flower organs and foliage leaves. It is perhaps not too difficult to imagine that sepals are equivalent to leaves because they usually have a very leaf-like appearance. Petals are also not so different from leaves, give or take a bit of shape and colour. But what about the sex organs? The male organs (stamens) do not bear any obvious resemblance to leaves. In the case of the female organs (carpels) we sometimes get a faint leaf-like appearance when they have been fertilised and grow into fruits or pods containing seeds: a pea pod could be thought of as a leaf that has been folded lengthways and had the edges stuck together. But what about a tomato? Slice a tomato cross-wise and you will see two or more segments, each containing seeds. Is a tomato several leaf-like organs joined together? The tomato segments do not look like leaves, so it is not at all obvious that they are the same sort of thing. As with his studies on the human skull, Goethe turned to abnormalities to help resolve the issue.

Helpful monsters
Monstrous flowers are curiously attractive. For years gardeners have selected varieties with extra petals, sometimes called double-flowered forms. Roses, for example, have only five petals in the wild, yet many of the commonly cultivated garden varieties have many more than this. They have been selectively bred for their appeal to humans. In some cases, these abnormal flowers have extra petals at the expense of sex organs, so they can no longer reproduce properly by sexual means (many of them are propagated vegetatively, by taking cuttings).
Although considered attractive to gardeners, most botanists viewed these abnormalities with suspicion, as unruly freaks of nature that would not repay further study. The eighteenth-century philosopher Jean Jacques Rousseau, also a keen botanist, warned young ladies against the dangers of such flowers:
Whenever you find them double, do not meddle with them, they are disfigured; or, if you please, dressed after our fashion: nature will no longer be found among them; she refuses to reproduce any thing from monsters thus mutilated: for if the more brilliant parts of the flower, namely the corolla [petals], be multiplied, it is at the expense of the more essential parts [sex organs], which disappear under this addition of brilliancy.
Rather than shunning these monstrosities, Goethe realised that they could provide important clues to understanding how flowers normally form. To Goethe, the monstrous flowers with extra petals in their centre suggested that the sex organs could somehow be transformed into petals. Surely this showed that the different organs of a flower were interconvertible and so fundamentally equivalent. If this conclusion was granted, then the obvious similarity between foliage leaves and at least some of the flower organs (sepals and petals) indicated that all of the organs of a plant should be lumped into the same equivalence group. The various parts of a flower were equivalent to each other and to other types of leaves; they were all variations on a common theme. As further confirmation of this idea, Goethe cited abnormal roses which, instead of sex organs, had an entire shoot emerging from their centre, bearing petals and leaves (Fig. 4.3). Here was a dear illustration of the equivalence between floral organs and leaves.
Fig. 4.3 Proliferating rose showing leafy shoot emerging from the centre
When Goethe wrote his essay on plant metamorphosis, he was not aware that some of the ideas had been arrived at twenty years before him, by Caspar Friedrich Wolff. Wolff was one of the founding fathers of the theory of epigenesis, the view that organisms develop by new formation rather than being preformed in the egg (Chapter 1). At the age of 26, Wolff had produced a doctoral dissertation at the University of Halle, Theoria Generationis, which was