

бх
.docxГосударственное учреждение «Днепропетровская медицинская академия» Министерства здравоохранения Украины
кафедра «Микробиологии, вирусологии, иммунологии и эпидемиологии»
Самостоятельная внеаудиторная работа
на тему «Мобильные генетические элементы прокариот»
студентки II курса, I медицинского факультета, группы 4-Б
Чайки Марии Вадимовны
Днепропетровск
2014 г.
Мигрирующие генетические элементы (мобильные гены, прыгающие гены), дискретные фрагменты (сегменты) ДНК, способные встраиваться в разные участки генома; их расположение на хромосомах может меняться как в процессе исторического развития мира организмов, так и в пределах жизни одного индивидуума.
Мобильные
гены открыты в 40-х гг. 20 в. Б.
Мак-Клинток
на основании генетического анализа
нестабильных мутаций у кукурузы.
Исследование их молекулярной природы
начато в 60-х гг. в связи с обнаружением
нового типа мутационных изменений у
бактерий (так называемых вставочных
мутаций) и идентификацией носителей
этих мутаций. Структурно-функциональные
исследования мгэ эукариот на молекулярном
уровне ведутся с кон. 70-х гг. с 
использованием методов клонирования (получение наследственно однородных поколений особи или клетки путем бесполого размножения) и генетической инженерии.
Существует несколько классов мобильных элементов генома, отличающихся по строению и способу перемещения:
-
Tn-транспозоны;
-
Is-инсерционные элементы;
-
ДНК-транспозоны;
-
Ретротранспозоны;
-
Плазмиды;
-
Бактериофаги;
-
Интроны второй группы.
У прокариот выделено несколько основных групп мгэ: плазмиды, инсерционные элементы и транспозоны, эписомы, а также некоторые бактериофаги.
Плазмиды - небольшие молекулы ДНК, физически отдельные от геномных хромосом и способные реплицироваться автономно.
Хромосомная ДНК (1) и плазмиды (2) в бактериальной клетке
Типы плазмид. Большинство плазмид классифицируют на основании тех свойств бактериальной клетки, которые привели к обнаружению этих плазмид:
1) F-факторы (fertility — плодовитость);
2) R-факторы (resistance — резистентность, устойчивость);
3) Соl-факторы (соlicinogeny — колициногенность);
4) пенициллиназные плазмиды золотистого стафилококка; 5) плазмиды деградации псевдомонад и др.
Плазмиды можно убрать (элиминировать) из бактерии нагреванием, акридиновыми красителями, ультрафиолетовыми лучами, подавляющими репликацию (воспроизведение) плазмиды. Удаление плазмиды не нарушает жизненно важные функции клетки.
F-факторы — плазмиды, которые определяют появление новых поверхностных структур клетки, — F-ворсинок, или пилей, позволяющих клеткам вступать в контакт (конъюгировать) и обеспечивать процесс переноса плазмидной ДНК из одной клетки в другую. Все плазмиды, которые сообщают своим хозяевам способность к переносу ДНК хромосомы, называют половыми.
R-фактор — плазмиды, которые обусловливают множественную резистентность микроорганизмов к лекарственным веществам. Впервые был обнаружен в Японии в 1955 г. во время вспышки дизентерии, при выделении штамма шигелл, устойчивых к четырем лекарственным препаратам: стрептомицину, тетрациклину, хлорамфениколу и сульфаниламиду.
R-фактор обычно находится в автономном состоянии в цитоплазме, но может встраиваться в хромосому и тогда выполняет функции полового фактора, обеспечивающего перенос хромосомы хозяина в другую клетку. Появление штаммов, устойчивых к антибиотикам и сульфаниламидным препаратам, затрудняет лечение инфекционных больных.
Соl-фактор, или фактор колициногенности, определяет способность бактерий образовывать особые вещества, которые вызывают гибель близкородственных штаммов.
Впервые эти вещества были обнаружены в культуре кишечной палочки, поэтому их назвали колицинами. Продукция веществ, подобных колицинам, в дальнейшем бы¬ла установлена и у других бактерий: холерного вибриона (вибриоцины), бактерий чумы (пестицины) и т. д. Эти вещества стали называть бактериоцинами. Они имеют белковую природу, обладают способностью адсорбироваться на поверхности бактериальной клетки, подавляют в ней обменные процессы и вызывают гибель клетки. Бактериоцины действуют только на бактерии, близкородственные продуценту. Продукция бактериоцинов чаще всего смертельна для клеток, продуцирующих их. Способность клетки к продукции бактериоцинов определяет автономная плазмида, называемая Соl-фактором. В естественных условиях только единичные клетки в популяции (1 на 1000) спонтанно продуцируют бактериоцины. При ультрафиолетовом облучении число продуцентов увеличивается/Способность бактериальных клеток продуцировать бактериоцины и специфичность их действия могут быть использованы для эпидемиологических целей при типировании культур, выделенных в очагах, с целью выявления источника инфекции. Предложена схема колицинотипирования возбудителей дизентерии.
Пенициллиназные плазмиды золотистого стафилококка обусловливают образование активного фермента пенициллиназы, который разрушает пенициллин. Поэтому антибиотик, эффективный в начале его применения при лечении стафилококковых инфекций, перестал оказывать действие на штаммы стафилококка, ставшие к нему устойчивыми.
|
Плазмида |
Хозяин |
Размер плазмиды (тыс. пар оснований) |
Геометрия плазмиды |
Число копий плазмиды на клетку |
|
pUB110 |
Bacillus subtilis |
2,3 |
Кольцевая |
20—50 |
|
ColEl |
Escherichia coli |
6,6 |
Кольцевая |
10—30 |
|
lp25 |
Borrelia burgdorferi |
24,2 |
Линейная |
1—2 |
|
pNOB8 |
Sulfolobus sp.a (архея) |
41,2 |
Кольцевая |
2—40 |
|
F |
Escherichia coli |
99,2 |
Кольцевая |
1—2 |
|
SCP1 |
Streptomyces coelicolor |
350,0 |
Линейная |
4 |
|
pSymA |
Sinorhizobium meliloti |
1354,2 |
Кольцевая |
2—3 |
|
Плазмида |
Хозяин |
Размер плазмиды (тыс. пар оснований) |
Известная функция |
|
pT181 |
Staphylococcus aureus |
4,4 |
Устойчивость к тетрациклину |
|
ColEl |
Escherichia coli |
6,6 |
Образование колицина и устойчивость к нему |
|
pMBl |
Escherichia coli |
8,5 |
Система рестрикции-модификации |
|
pGKL2 |
Kluyveromyces lactisb |
13,5 |
Плазмида-киллер |
|
pAMpi |
Enterococcus faecalis |
26,0 |
Устойчивость к эритромицину |
|
pSK41 |
Staphylococcus aureus |
46,4 |
Множественная устойчивость |
|
pBM4000 |
Bacillus megaterium |
53,0 |
Оперон рРНК |
|
pI258 |
Staphylococcus aureus |
28,0 |
Устойчивость к ионам тяжёлых металлов |
|
pSLT |
Salmonella enterica ssp. typhimurium |
93,9 |
Детерминанта вирулентности |
|
pMT1 |
Yersinia pestis |
101,0 |
Детерминанта вирулентности |
|
pADP-1 |
Pseudomonas sp. |
108,8 |
Катаболизм атразина (гербицид) |
|
pWW0 |
Pseudomonas putida |
117,0 |
Деградация ароматических углеводородов |
|
pX01 |
Bacillus anthracis |
181,7 |
Синтез энтеротоксинов |
|
pSOL1 |
Clostridium acetobutylicum |
192,0 |
Образование сольвента |
|
pSymB |
Sinorhizobium meliloti |
1683,3 |
Множественные функции |
IS-элементы - простые вставочные (инсерционные) последовательности, они обозначаются в зависимости от их нуклеотидного состава номерами IS1, IS2 и т.д., содержат от 700 до 1500 пар нуклеотидов. Эти сегменты ДНК имеют инвертированные повторы на концах, содержащие обычно несколько десятков нуклеотидных пар, и не содержат никаких генов, кроме тех, которые необходимы для их перемещения (транспозиции) по геному. Они встречаются в некоторых плазмидах (внехромосомные носители наследственности) и умеренных фагах (способны существовать в клетке в форме профага). Так, у разных штаммов бактерии Escherichia coli (E. coli) присутствует в геноме 19 копий IS1-элементов. Большинство др. IS-элементов также представлено в хромосомах разных штаммов E. coli многочисленными копиями: IS2-от 0 до 12,IS3-от 4 до 6, IS4-от 1 до 2, IS5-от 0 до 10.
Обычно IS-элементы встраиваются (интегрируют) в различные места бактериального генома, однако некоторые участки оказываются более предпочтительными, чем другие. Встраивание и исключение этих элементов происходит с высокой точностью, что свидетельствует об участии в этих процессах ферментов, узнающих инвертированные концевые повторы IS-элементов.
Ферментные системы, обусловливающие транспозиции IS-элементов, по крайней мере, частично кодируются их собств. ДНК. Так, IS1, судя пo длине его нуклеотидной последовательности, может кодировать лишь небольшие полипептиды. которые участвуют в его транспозиции, вероятно, в комплексе с клеточными белками. Значение IS-элементов для эволюции бактерий связано с тем, что эти элементы при своих перемещениях инактивируют различные гены или нарушают их нормальную регуляцию. Помимо прямого влияния на экспрессию гена (развития признака, контролируемого данным геном) вследствие транспозиции инсерционной последовательности непосредственно в кодирующую часть гена или его регуляторную зону, эти мгэ могут влиять также на транскрипцию окружающих их последовательностей ДНК генома. Это происходит вследствие того, что многие IS-элементы содержат промоторные и терминаторные участки ДНК. Транспозиции IS-элементов могут вызывать слияние двух не связанных ранее генов или оперонов (совокупность связанных между собой генов и прилегающих к ним регуляторных участков) с образованием новых функционых единиц, а также индуцировать все виды хромосомных перестроек. Соединение разнородных репликонов (элементарная генетическая структура, способная к самокопированию) имеет большое биологическое значение, т. к. объединяет ранее разобщенные генетические детерминанты, подчас принадлежащие разным видам организмов. Tn-элементы (сложные перемещающиеся элементы, или транспозоны) принципиально отличаются от IS-элементов только тем, что содержат дополнит. структурные гены, не имеющие отношения к ф-ции транспозиции. Известно много транспозонов, в состав которых входят гены устойчивости к антибиотикам, тяжелым металлам и другим ядам. При этом один и тот же транспозон иногда несет целый набор Детерминант резистентности (т. наз. V-детерминанты). Такие транспозоны наиболее широко распространены, т.к. представляют ценность для селекции бактерий. Существуют транспозоны, содержащие гены, которые кодируют токсины, а также свойственные данному организму ферменты. Как правило, Tn-элементы несут на концах целые или частично измененные IS-элементы, которые сообщают им способность перемещаться по геному и вызывать в нем те же изменения, что и своб. IS-элементы. При этом 2 концевые IS-подобные терминальные последовательности в зависимости от типа транспозона могут иметь прямую или инвертир. последовательность нуклеотидов. Разные транспозоны часто содержат одинаковые терминальные последовательности нуклеотидов.
Структура
генома типичного автономного ДНК
транспозона

ITR – инвертированные повторы;
Transposase – ДНК эндонуклеаза - ДНК лигаза;
DR – дупликация сайта встраивания.
Транспозоны вместе с плазмидами и фагами (в которые они легко интегрируются) способны осуществлять обмен различных заключенных в них генов между весьма отдаленными видами бактерий, поэтому они играют чрезвычайно важную роль в эволюции бактерий, включая адаптацию их к лек. веществам и продуцирования ими новых токсинов.
Транспозиция Tn-элементов осуществляется по такому же механизму, как и IS-элементов, и также включает стадию трансляции. Большинство транспозонов не выбирает для своего включения строго определенные последовательности в ДНК. Однако обычно они предпочитают некоторые районы хромосом и даже специфические участки, причем разные Тn-элементы различаются по специфичности выбора мест интеграции.
Частота и характер перемещений IS- и Тn-элементов варьируют в весьма широких пределах и зависят прежде всего от свойств самих элементов. Например, Тn-З плазмиды перемещаются чаще в др. плазмиды, чем в хромосому. На транспозиции влияют не только генетич., но и различные внешние факторы, например УФ облучение. По-видимому, яды, инактивация которых обусловлена генами транспозонов, могут индуцировать синтез ферментов, необходимых для транспозиции этих транспозонов.
-
Транспозаза дважды надрезает только одну нить ДНК в спирали.
-
В месте нового встраивания она так же создаёт один надрез.
-
Транспозаза переносит концы нити ДНК транспозона в место надреза и лигирует с ДНК хозяина.
-
Репликация создаёт копию транспозона по двум нитям на новом и старом месте.

Другую группу мгэ бактерий составляют эписомы - сложные плазмиды, способные к интеграции в хромосому. Эписомы, как правило, содержат IS- или Tn-элементы, и в большинстве случаев именно благодаря им они могут включаться в состав хромосомы. Так, в половой F-эписоме E. coli (мол. м. 6.107) имеется одна копия IS2, две копии IS3 и одна копия Тn1000.
К
мгэ прокариот относят также умеренные
фаги.
l-Фаги (лямбдоидные фаги) обычно
встраиваются в одно место хромосомы,
но при определенных условиях могут
располагаться и в др. участках генома.
m-Фаги способны включаться в любые места
бактериальной хромосомы, а также в ДНК
мн. др. фагов и плазмид. Интеграция
лямбдоидных фагов обеспечивается
ферментной системой, состоящей из
клеточных белков и белков, кодируемых
геномом фага. m-Фаг во мн. отношениях
сходен с IS- и Tn-элементами и отличается
от них только тем, что может формировать
вирусные частицы. Предполагают, что IS-
и Тn-элементы произошли из фага типа C
в результате утери большинства его
генов.

Умеренные фаги способны вносить существ. изменения в структуру и функционирование бактериального генома благодаря двум процессам - интеграции фаговой ДНК в хромосому бактерии и трансдукции (переносу фагом бактериальных генов из одних клеток в другие). Трансдуцирующие фаги образуются в результате неточного исключения из хромосомы интегрированной фаговой ДНК. При этом часть собственной ДНК фага утрачивается, и вместо нее в фаговый геном включается участок бактериальной ДНК, достигающий иногда значительных размеров. Интегрированные фаги могут мутировать и терять способность к исключению из хромосомы, становясь вследствие этого ее неотъемлемой частью. В этом случае гены фага начинают определять функции клетки, т.е. становятся ее собственными генами.
Стратегии жизненных циклов мобильных элементов основаны на использовании толерантности клетки к ДНК уже имеющей связь с клеточной ДНК.
-
Частично интегрированную ДНК клетка считает своей и «корректирует» разрывы системой репарации. Но в случае, если репликация ДНК и обратная транскрипция происходят в цитоплазме, они являются объектом атаки защитных комплексов клетки на цитоплазматическую ДНК и ДНК-РНК гибриды. Это обуславливает необходимость наличия изолирующего «вирусного» протеинового капсида, проницаемого для простых соединений и не проницаемого для крупных молекул.
-
Мобильные элементы создают большое количество собственных копий. Транспозиции IS-элементов не сопряжены с их исключением из мест исходной локализации в плазмидах или хромосоме; при транспозиции IS-элемент удваивается и одна его копия остается на прежнем месте, а другая попадает в новый локус (местоположение гена в хромосоме или плазмиде). Таким образом, транспозиции этого элемента сопряжены с репликацией (удвоением) его ДНК. Создание множества своих копий, с другой стороны, делает возможным для клетки распознавание и нейтрализацию активности мобильных элементов.
-
TPRT (обратная транскрипция) является более защищенной и простой реакцией, однако подвержена «смене матрицы» и для полноценного воспроизведения копий требует наличия элементов внутреннего промотора элемента. Интроны могут так же быть удобны для воспроизводства элементов, не имеющих рабочего внутреннего промотора.
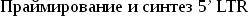
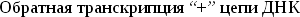

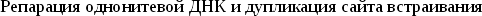
-
Среди мобильных элементов не является редкостью чувствительность к последовательности сайта встраивания. Однако, как правило, эти последовательности достаточно просты и встречаются в геноме многократно.
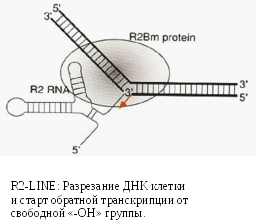
-
Как автономные, так и не автономные мобильные элементы мимикрируют, подстраиваясь под существующие в клетке структуры.
Неавтономный
SINE-элемент

-
Многокопийные функциональные некодирующие короткие РНК (такие как тРНК) часто становятся основой не автономных элементов, ввиду толерантности клетки к числу копий этих РНК, наличию у них элементов внутреннего промотора и способности взаимодействовать с протеиновыми клеточными структурам.
-
Важным отличием мгэ эукариот от таковых у бактерий является их способность при включении в тот или иной локус изменять свойства ферментов (продуктов генов-мишеней), а не только прерывать их синтез.
|
Процесс |
Какие элементы используют |
|
Транскрипция |
Все автономные и все ретро-элементы. |
|
Трансляция |
Все автономные элементы. |
|
Вырезание ДНК |
Все транспозоны. |
|
Встраивание ДНК |
Все транспозоны, LTR элементы, DIRS. |
|
Обратная транскрипция |
Все ретроэлементы. |
|
Репликация кольцевой ДНК |
Некоторые ретровирусы, DIRS, Полинтоны. |
Хотя мобильные элементы в целом являются «генетическими паразитами», вызывая мутации в генетическом материале организма хозяина и понижая его приспособленность за счёт траты энергии на репликацию и синтез белков паразита, они являются важным механизмом изменчивости и обмена генетическим материалом между организмами одного вида и разными видами.