палеонтология бондаренко
.pdfМОСКОВСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ им. MB. ЛОМОНОСОВА
ПАЛЕОНТОЛОГИЧЕСКИЙ ИНСТИТУТ РАН ФОНД ХАНСА РАУСИНГА
И.А. Михайлова, О.Б. Бондаренко
ПАЛЕОНТОЛОГИЯ
ЧАСТЬ 1
Рекомендовано Министерством общего и профессионального образования Российской Федерации в качестве учебника для студентов высших учебных заведений, обучающихся по направлению и специальности «Геология»
850-летию Москвы посвящается
. V
ИЗДАТЕЛЬСТВО МОСКОВСКОГО УНИВЕРСИТЕТА 1997
ББК 28.1
М69 УДК 56 (075.8)
Р е ц е н з е н т ы :
кафедра региональной геологии и палеонтологии МГГА; доктор биологических наук, профессор В.Н. Шиманский
БИБЛИОТЕКИ |
Сыктывкарского {
ГОСУНИЗЕРСИТЕТЯ1
Михайлова И.А., Бондаренко О.Б.
М69 Палеонтология. Ч. 1: Учебник. —. М.: Изд-во МГУ,, 1997. — 448 с: ил.
ISBN 5-211-03841-Х (Ч. 1) ISBN 5-211-03868-1
В первой части учебника изложены основные сведения об ископае мых бактериях, цианобионтах, растениях, грибах и животных (беспозво ночных и позвоночных). Описана геологическая история как отдельных групп, так и органического мира в целом. Рассмотрены общие вопросы: история становления и разделы палеонтологии, закономерности эволю ции, биотические события и т.д. Учебник проиллюстрирован много численными рисунками (более 250), в значительной степени новыми и оригинальными
Для студентов геологических, географических и биологических факуль
тетов университетов и других вузов, а также специалистов палеонтологов.
ББК 28.1
ISBN 5-211-03841-Х (/% 1^ |
|
ISBN 5-211-03868-1 |
© Михайлова И.А., Бондаренко О.Б., 1997 г. |
П Р Е Д И С Л О В И Е
После выхода в свет учебника «Общая палеонтология» про шло более семи лет. Приступая к работе над новой книгой, авто ры старались учесть все замечания и новые данные. Исключи тельную помощь оказали официальные и неофициальные рецен зенты, благодаря которым произошла радикальная переработка разделов, посвященных ископаемым растениям и типу хордовых. Мы не питаем иллюзий, что нам удалось полностью избежать до садных неточностей и упущений, но искренне надеемся, что они сведены до минимума.
При создании учебника авторы стремились максимально про иллюстрировать текст, исходя из тезиса «лучше один раз увидеть...». Следует также иметь в виду, что рисунки (их более 250) по содер^ жанию намного информативнее и компактнее, чем текст. Допол нительными иллюстрациями являются четыре форзаца. На пер вом из них приведена циклограмма процентного содержания ви дов различных типов царства животных, а внутри каждого типа показано соотношение современных и ископаемых форм (Пале онтология беспозвоночных, 1962). Форзац 2 посвящен распреде лению водных организмов в зависимости от солености бассейна (Fenton, Vickers-Rich, Rich, 1989). На форзаце 3 приведены четы ре отличающиеся реконструкции рода Iguanodon (динозавр), пред ложенные разными авторами. Форзац 4 иллюстрирует развитие органического мира на протяжении геологической истории Земли.
Учебник состоит из трех частей: вводной, систематической и заключительной. Вводная часть содержит представления о типах сохранности, химическом составе и породообразующей роли ис копаемых; историю становления и разделы палеонтологии; све дения об условиях обитания и образе жизни; рассмотрение зако номерностей эволюции; анализ биотических событий с точки зре ния возникновения жизни, массовых появлений и массовых вымираний организмов крупного таксономического ранга. В конце вводной части приведена Международная геохронологическая (стратиграфическая) шкала.
1* |
3 |
Систематическая часть начинается с изложения общих по ложений, касающихся классификации, таксономии и системати ки, а также объяснения процедурных приемов обозначения так сонов и особенностей их применения в палеонтологии. Основ ной объем систематической части, как и учебника в целом, посвящен характеристикам конкретных систематических единиц (морфология, классификация, условия обитания и образ жизни, породообразующая роль, геологический возраст, история разви тия). Приведены диагнозы следующих подразделений (в скобках указано число единиц): надцарств (2), царств (5), подцарств (4), надразделов и надотделов (4), разделов (2), подразделов (2), ти пов и отделов (29), классов (61), отрядов и подотрядов (113), не считая диагнозов подтипов, подклассов и надотрядов.
В заключительной части рассмотрена история органического мира по зонам и периодам. Учебник завершает список рекомен дуемой литературы и указатель латинских названий.
Учебник подготовлен сотрудниками кафедры палеонтологии геологического факультета Московского государственного универ ситета И.А. Михайловой и О.Б. Бондаренко. Самую искреннюю признательность авторы хотят выразить директору Палеонтоло гического института Российской Академии наук профессору еА.Ю. Розанову, без действенной помощи которого эта работа могла
еще долго ждать опубликования. А.Ю. Розанов знакомился с сожанием учебника в процессе его создания и сделал существен ные замечания по Международной геохронологической (стратиг рафической) шкале, а также внес много нового в раздел, посвя щенный археоциатам. Благодаря настойчивым усилиям дирекции ПИН РАН была получена финансовая поддержка со стороны фонда Ханса Раусинга. Авторам особенно приятно, что учебник
выходит в свет в год 850-летия г. Москвы.
При подготовке рукописи авторы постоянно консультирова лись с коллегами кафедры палеонтологии МГУ, Палеонтологи ческого и Геологического институтов РАН. Это также официаль ные рецензенты Т.Т. Шаркова, Г.Н. Садовников (МГГА) и В.Н. Шиманский (ПИН РАН). Для коренной переработки палео ботанического раздела важнейшее значение имели обсуждения дис куссионных положений с Г.Н. Садовниковым. Раздел, посвящен ный позвоночным, был внимательно прочитан В.И. Жегалло и улучшен за счет предоставления им новых материалов и разъяс нения спорных вопросов. О.П. Обручева, являвшаяся нашим со автором в предыдущем учебнике «Общая палеонтология», про смотрела готовый материал по позвоночным и сделала некоторые замечания. Мы выражаем искреннюю признательность и благо дарность всем тем, кто в той или иной мере способствовал улуч шению содержания учебника.
Особо хотим отметить роль студентов и в первую очередь сту дентов палеонтологов. Их насмешливый и критический ум заста вил авторов более аккуратно отнестись к изложению и определе нию ряда терминов, за что мы им искренне благодарны. К величай шему сожалению, жесткие рамки объема учебника препятствовали нашему стремлению изложить то или иное положение более под робно, а также внести некоторые дополнительные сведения. За помощь и консультации по изданию книги мы признательны вы пускнику нашей кафедры А.В. Вараксину.
В создании учебника с самого начала активно участвовала М.В. Кнорина, постоянно помогавшая авторам избегать ошибок, разночтений и повторов; ею составлен указатель латинских на званий. Большая часть иллюстраций подготовлена сотрудниками картбюро геологического факультета МГУ под руководством А.Г. Дубровина, причем значительная часть рисунков была вы полнена Е.П. Загозиной. Авторы им глубоко признательны и благодарны.
4
ВВОДНАЯ ЧАСТЬ
Палеонтология изучает органический мир прошлого. Столь удачное название «палеонтология» (греч. palaios — древний; on, ontos — существо; logos — понятие, учение), хорошо отражающее суть науки о древних существах, предложил в 1822 г. французс кий зоолог А.М.Д. де Блэнвилль (1777-1850). Вскоре, в 1834 г., профессор Московского университета Г.И. Фишер фон Вальдгейм принял этот термин, хотя ранее он использовал название «петроматогнозия». Более того, он не только сам стал употреблять тер мин «палеонтология», но и всячески способствовал его внедрению
внаучную и учебную литературу. Поэтому Г.И. Фишера наравне
сА.М.Д. де Блэнвиллем считают автором термина «палеонтология».
Объектами палеонтологии являются ископаемые, т.е. те орга нические остатки, по которым мы можем судить о животных и растениях прошлых геологических эпох, а также о бактериях, цианобионтах и грибах, некогда живших на нашей планете. Поэто му палеонтология входит в цикл биологических дисциплин и для нее вполне применимо название «палеобиология».
Цель палеонтологии как биологической науки — воссоздание органического мира прошлого с его законами развития но време ни и в пространстве, а одна из основных практических задач — установление относительного возраста отложений по комплексам ископаемых остатков.
ТИПЫ СОХРАННОСТИ ИСКОПАЕМЫХ
Объектами палеонтологических исследовании являются лю бые ископаемые биологического происхождении К ним относят ся не только сами организмы, но и биогеоческие компонен ты, возникшие в биосферах прошлого при участииорганизмов. Для всех них широко используют термиы органические остат ки», «ископаемые», «окаменелости» и "фоссилии" fossilis — погребенный, ископаемый). Последний термин в настоящее вре мя очень популярен в зарубежной и русскоязычной литературе, хотя его использовали уже во времена Екатерины II.
6
Фоссилизация. Процессы преобразования погибших организ мов в ископаемые называют фоссилизацией. Гибель и последую щее захоронение организмов сопровождается воздействием раз личных факторов среды. Они проходят все процессы диагенеза, т.е. физических и химических преобразований при переходе осадка в породу, в которую они заключены.
После гибели организма в первую очередь разрушаются мяг кие ткани, затем начинается заполнение пустот скелета вмещаю щим осадком или минеральными соединениями. Иногда пустоты скелета подвергаются пиритизации, ожелезнению, часто в них возникают друзы и щетки кальцита, аметиста, флюорита, галени та и т.д. Ископаемые скелеты нередко оказываются заключенны ми в фосфоритовые конкреции. При фоссилизации скелеты под вергаются перекристаллизации, приводящей к более устойчивым минеральным модификациям. Например, арагонитовые раковин ки преобразуются в кальцитовые. Нередки случаи минерализации, когда первичный химический состав скелета изменяется (псевдо морфозы). Так, известковые раковины частично или полностью замещаются водным кремнеземом и наоборот. Также наблюдают ся фосфатизация, пиритизация и ожелезнение минеральных и органических скелетов.
Растения при фоссилизации обычно разрушаются полностью, чему способствуют процессы гниения и брожения. Тем не менее ископаемые растения обнаружены начиная с докембрия. Чаще всего от растений сохраняются обугленные остатки листьев, листоподобных образований, стеблей, стволов, корней, семян, пло дов, шишек, спор и пыльцы. В процессе фоссилизации первич ные растительные ткани могут разрушаться полностью, и тогда остаются отпечатки и ядра. Нередко при фоссилизации расти тельные ткани замещаются различными минеральными соедине ниями, чаще всего кремнеземом, карбонатом и пиритом. Подо бное полное или частичное замещение стволов растений при со хранении внутренней структуры называется петрификация. Чаще всего окаменение стволов связано с замещением кремнеземом, реже пиритом, кальцитом и др. Известны скопления стволов, так называемые окаменелые леса. Это либо целые стволы, либо толь ко их нижние части.
Некоторые органические образования растений (воск, смола, лигнин, целлюлоза) сохраняются в ископаемом состоянии, почти не изменяясь. Минеральные слабоизмененные компоненты рас тений встречаются также довольно часто: это кремневые рако винки диатомовых водорослей, известковые «плодовые шарики» харовых растений, известковые пленки и желваки красных водо рослей и т.д.
7
В зависимости от полноты сохранности и своеобразия остат ков выделяют следующие категории ископаемых: субфоссилии, эуфоссилии, ихнофоссилии, копрофоссилии, хемофоссилии.
Субфоссилии (лат. sub — под, почти) представлены ископае мыми (почти ископаемыми), у которых сохранился не только ске лет, но и слабоизмененные мягкие ткани. Для растительных ос татков используют термин «фитолеймы» (греч. phyton — расте ние; leimma — остаток). Это в различной степени измененные растительные остатки, сохраняющие клеточную структуру. К субфоссилиям относятся фитолеймы из четвертичных отложений — семена, орехи, шишки хвойных, древесина, захороненные в тор фяниках. Более измененные фитолеймы являются эуфоссилиями. К субфоссилиям принадлежат и уникальные находки некото рых животных этого времени, например мамонты, носороги и птицы. Консервантами для таких ископаемых являются вечная мерзлота, различные битумы, вулканические пеплы, эоловые пес ки. Считалось, что и янтарь является превосходным консерван том, однако в янтаре не сохраняются мягкие ткани. Вместе с тем ископаемые в янтаре (и растения и животные) полностью сохра няют свою форму, что позволяет тщательно изучить их внешнюю морфологию. Но попытка извлечь объект заканчивается тем, что все содержимое рассыпается в пыль. Значительно реже субфосси лии встречаются в отложениях более древних, чем четвертичные.
Эуфоссилии, или эвфоссилии (греч. ей — хорошо, настоящий) представлены целыми скелетами или фрагментами скелетов и их дискретными элементами, а также отпечатками и ядрами. Скеле ты являются основными объектами палеонтологических исследо ваний. Скелетные остатки имеют минеральный или органичес кий состав. Это раковины и скелеты животных, оболочки бакте рий и грибов, а также органические остатки листьев, семян, плодов, спор и пыльцы. Особо следует сказать о фитолеймах, представ ленных в различной степени обугленными остатками листьев, древесины, семян, плодов, спор и пыльцы.
Иногда используется термин «органикостенные микрофосси лии», к которым относятся оболочки бактерий и грибов, нитча тых цианобионтов, а также споры и пыльца. Размеры таких фоссилий менее 100 мкм. Органикостенные микрофоссилии могут иметь очень хорошую сохранность даже в докембрийских отло жениях (см. рис. 250). Термин «органикостенные» можно распрос транять и на макроостатки.
Многие эуфоссилии сохраняют информацию не только о мяг ких частях организма и его функциональных системах, таких как кровеносная, половая, проводящие пучки растений и др., но и об образе жизни и биогеохимических процессах.
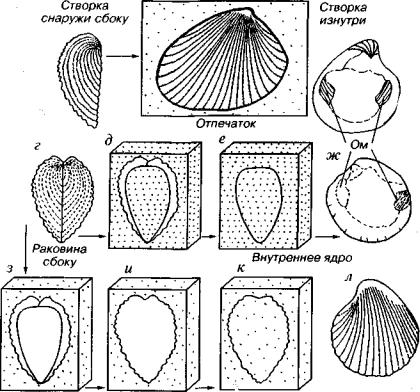
От скелетов и мягких частей организмов могут сохраняться отпечатки и ядра. Отпечатки представляют собой уплощенные оттиски, а ядра — объемные слепки полостей (рис. 1). Некоторые животные известны только по отпечаткам. Наиболее знамениты ми являются местонахождения отпечатков птиц, рыб, медузоидов, червей, членистоногих и других животных, найденные в юр ских золенгофенских сланцах Германии и в вендских отложениях Эдиакары Австралии. От растений чаще всего встречаются отпе чатки листьев, реже стволов, семян и др. Отпечатки листьев отра жают не только форму, но и характер жилкования. Отпечатки стволов сохраняют особенности поверхностного строения коры, например листовые подушки лепидодендроновых.
Среди ядер различают внутренние и внешние. Внутренние ядра возникают за счет заполнения породой внутренних полостей
а |
б |
в |
Внешнее ядро
Рис. 1. Схема образования отпечатка (а-б), внутреннего ядра (в-ж) и внешнего ядра (з-л) на примере ребристой раковины двустворок. Ом — отпечатки мускулов
К |
•1 -2697 |
Q |
|
|
раковин двустворок, остракод, гастропод, брахиопод, аммонитов, а также черепных коробок позвоночных животных. Ядра расте ний чаще всего представляют отливы сердцевины стволов. Про цесс возникновения внешних ядер сложнее, чем внутренних. Сна чала скелет, заключенный в породе и ограничивающий полость, растворяется. Затем начинается заполнение породой вновь воз никшей полости. Внутренние и внешние ядра наиболее четко от личаются друг от друга у скульптированных двустворок и брахио под На внутреннем ядре имеются отпечатки различных внутрен них структур, а наружное ядро отражает особенности скульптуры раковины. Внешние ядра ребристые, шероховатые, грубые, а внут ренние — гладкие, с отпечатками мускулов, связок и других эле ментов внутреннего строения.
Ихнофоссшши (греч. ichnos - след) представлены следами жизнедеятельности ископаемых организмов. Чаще всего они со храняются в виде отпечатков, реже в виде слабообъемных образо ваний. К ихнофоссилиям относят следы ползания и зарывания членистоногих, червей, двустворок; следы выедания, норки, ходы и следы сверления губок, двустворок, членистоногих; следы пе редвижения позвоночных.
Копрофоссилии (греч. kopros - помёт, навоз) состоят из продук тов жизнедеятельности ископаемых организмов. Они имеют объ емный характер, сохраняясь в виде валиков, желваков, конкреции, холмиков столбиков и даже пластовых тел. Термин «копрофосси лии» был предложен в 1989 г. в учебнике «Общая палеонтология». За основу взято название «копролиты», введенное в научную ли тературу свыше 150 лет назад и обозначающее «окаменевшие эк скременты животных» (Палеонтологический словарь, 1965).
К наиболее типичным копрофоссилиям относятся конечные продукты пищеварения илоедов и позвоночных животных; во вто рой группе копрофоссилии могут сохраняться непереваренные ос татки других животных и растений. Копрофоссилии илоедов пред ставлены валиками и ленточками, которые, на первый взгляд, как будто не отличаются от окружающей породы. Но, пройдя через кишечник илоеда, осадок обогащается кальцием, железом, магни ем, калием и фосфором. В результате копрофоссилии илоедов при обретают более светлый или, наоборот, более темный, нередко крас новатый оттенок, что и отличает их от окружающей породы. Про цесс переработки осадка илоедами и биофильтра юрами называют биотурбацией. Большинство оглж.чпмх оможеипй настоящего и прошлого проходят биотурбапит К..и.-тыг щюлукты биотурбации и следы жизнедеятельное ж mmi/u M.I u.in.ii«.i опт лифами.
К копрофоссилиям мож! |
1 |
к HIHIIIVKM.I жизнедеятель |
ности бактерий и циапобио.. |
I-.. |
и !•.... принимают участие в |
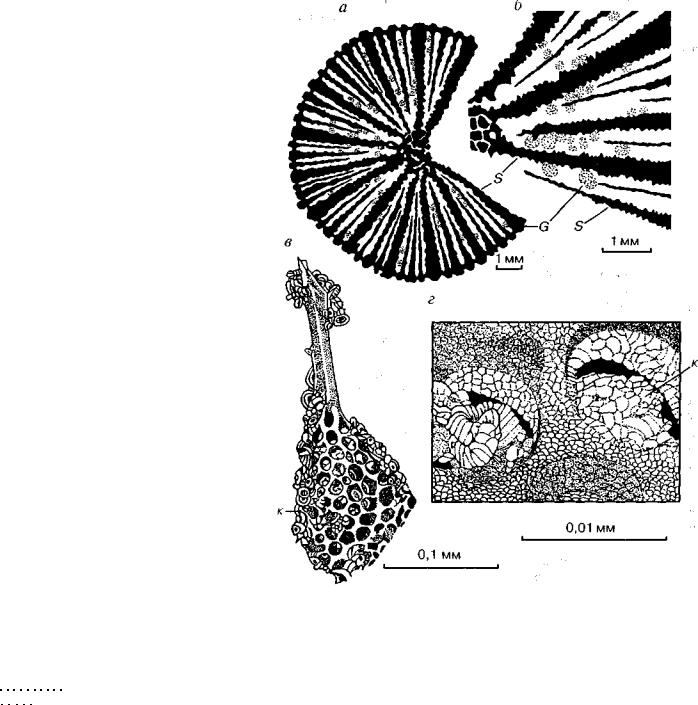
Рис. 2. Соотношения размеров скелетов различных групп организмов с;, б — между септами коралла (макрофоссилии) застряли раковинки форамини-
фер (микрофоссилии); в-г — в «окошечках» скелета радиолярии (микрофоссилии) находятся кокколиты (нанофоссилии). G — Globigerinida, к — кокколиты, S — септы
2*
10
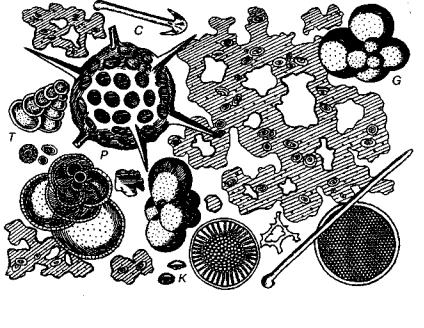
Рис. 3. Глубоководный органогенный ил (увел, в 700 раз), видна слизь с заклю ченными в ней кокколитами (К), раковинки Globigerinida (G) и Textulariida (Т), а также радиолярии (Р) и спикулы губок (С) (Циттель, 1934)
образовании железистых, марганцевых и фосфоритовых конкре ций, графитов, серы, нефти, газа и т.д. От жизнедеятельности цианобионтов сохраняются известковые слоистые образования — строматолиты, онколиты и катаграфии.
К хемофоссилиям (греч. chemie — химия) относят органичес кие ископаемые биомолекулы бактериального, цианобионтного, растительного и животного происхождения. Хемофоссилии со храняют химический состав биомолекул, позволяющий опреде лить систематическое положение исходного организма, но не его морфологию. Изучение химического и таксономического разно образия хемофоссилии тесно связано с проблемами возникнове ния и развития жизни, а также с происхождением горючих иско паемых, особенно нефти. Биологический фактор в формирова нии нефти долгое время отрицали, считая ее только хемогенной. Успехи в изучении хемофоссилии доказывают обратное. Хемо фоссилии являются объектом изучения биохимии и молекуляр ной палеонтологии.
Взависимости от размеров ископаемых можно выделить (рис. 2
и3): макрофоссилии (более 1 мм), микрофоссилии (десятые и сотые доли миллиметра) и нанофоссилии (сотые доли миллимет ра и менее).
12
За фоссилии, кроме перечисленных форм сохранности, иногда принимают различные минералогические и литологические об разования, получившие название псевдофоссилии (греч. pseudos — ложь). Разрозненные игольчатые минералы ошибочно описыва ют как спикулы губок, лучистые минеральные агрегаты — как колониальные кораллы, марганцевые дендриты — как растения, округлые желваки осадочного происхождения — как медузы и т.д.
ХИМИЧЕСКИЙ СОСТАВ И ПОРОДООБРАЗУЮЩАЯ РОЛЬ ИСКОПАЕМЫХ
Породообразующими называются ископаемые, которые со ставляют 30-40% и более от общего объема отложений. В образо вании органогенной породы принимают участие как скелетные остатки, так и продукты жизнедеятельности, т.е. эуфоссилии и копрофоссилии. Иногда такие породы называют биолитами. Не пременным условием породообразования является «скученный» характер обитания организмов. Таким свойством обладают в ос новном прикрепленные, малоподвижные и зарывающиеся фор мы, образующие заросли, банки, «луга», рифы и любые другие массовые поселения. Уже при жизни подобные организмы со ставляют основную часть биоценоза. Среди живущих в толще воды планктонных и нектонных организмов породообразующую роль играют те, которые обитают «стаями». Для континентальных ор ганизмов необходимы те же предпосылки. Органогенные породы могут возникать и за счет переноса первично единичных скеле тов, образующих вторичные скопления в различных понижениях рельефа.
Состав скелетов ископаемых может быть минеральным, ми нерально-органическим и органическим. Среди минеральных ске летов наиболее распространены известковые, кремневые и фос фатные, образующие соответствующие по составу породы. Осо бенно много органогенных пород известкового (= карбонатного) состава (СаСОэ): известняки, мергели, писчий мел, первичные и вторичные доломиты. Для названия органогенных пород, в отли чие от хемогенных, используют прилагательное от латинского (гре ческого) названия той группы организмов, которая является ос новным породообразователем. Среди органогенных известняков и мраморов, образованных скелетами животных, наиболее рас пространены фораминиферовые, коралловые, строматопоровые, археоциатовые, губковые, серпулитовые, гастроподовые (птероподовые, спирателловые и др.), цефалоподовые, остракодовые, мшанковые, брахиоподовые и криноидные. Известковые породы,
13
состоящие из скоплений раковинок двустворок, обычно называ ют ракушечниками (ракушняками), устричными горизонтами, рудистовыми известняками.
Скопления костей и скелетов позвоночных называют ней трально — «костеносные слои». Растения, имеющие карбонатные скелеты, дают начало известнякам — водорослевым, литотамниевым, харовым и кокколитовым (писчий мел). Органогенные из вестняки могут возникать и как конечные продукты жизнедея тельности цианобионтов и бактерий. От них остаются слоистые пластовые, желваковые, столбчатые, концентрические и «фигур ные» образования — строматолиты, онколиты и катаграфии.
Для скоплений измельченных известковых скелетов и рако вин различных систематических групп используют название «ор ганогенный детрит». Обугленные остатки растений неопредели мого систематического состава называют растительным детритом.
Минеральные скелеты кремневого (опалового — Si02 nH2 0) состава встречаются реже, чем карбонатного. Они известны у од ноклеточных животных (радиолярии, солнечники), у многокле точных примитивных животных (губки), а также у низших од ноклеточных водорослей (диатомовые и кремневые жгутиковые). Они образуют биогенные силициты. Кремневые породы — ра диоляриты — состоят из скелетиков радиолярий, спонголиты — из спикул губок, диатомиты — из створок диатомовых водоро слей. В каждой из перечисленных пород могут принимать участие кремневые скелеты других групп организмов. Так, в диатомитах иногда встречается небольшое количество радиолярий и спикул губок. А все они могут встречаться в таких биогенно-хемогенных породах, как трепел и опока.
Фосфатные скелеты в чистом виде встречаются редко, но фос фаты кальция (СаР04 — в простейшем случае) как примесь или основная составляющая известны у многих организмов. Фосфаты кальция обнаружены у бактерий, грибов, книдарий (статолиты медуз и оболочки конулярий), червей, членистоногих, моллюсков, брахиопод, мшанок, иглокожих и позвоночных (конодонты, зубы акул, зубы и кости млекопитающих). Благодаря концентрации биоген ного фосфата возникают месторождения фосфоритов. В Эстонии, например, источником биогенного фосфорита являются фосфори- то-хитиновые раковины брахиопод ордовикского рода Obolus s.l. (оболовые песчаники). В Подмосковье (например, в Лопатинском карьере) центрами фосфоритизации являются раковины позднеюрских аммоноидей. Фосфатная составляющая сконцентрирована в виде конкреций, желваков, оолитов и пластовых тел.
Минеральные скелеты всегда имеют органический матрикс, а также содержат элементы-примеси К, Mg, Ва, Si, Fe, Mn, Na, Ti,
14
F, Zn, Pb, Cu, Sr и др. В настоящее время обнаружены около 40 минералов, входящих в состав различных скелетов (табл. 1).
Кроме известковых и кремневых скелетов известны и такие экзотические, как целестиновые (SrS04), встречающиеся у акантарий (одноклеточные). Несмотря на то что при жизни акантарии образуют скопления, они не стали породообразующими, так как их игольчатые скелетики после отмирания растворяются в воде, не попадая в осадок.
За счет жизнедеятельности бактерий образуются железистые, марганцевые, медистые и сульфидные месторождения, такие как железистые кварциты (джеспилиты) Кривого Рога, медистые пес чаники Джезказгана. Бактерии участвуют в накоплении бокситов и фосфоритов.
Органические скелеты построены в основном углеводами, бел ками и жирами. Среди органических скелетов наиболее распрос транены хитиновые, хитиноподобные, шелкоподобные (спонгин), роговые и целлюлозные. Они образуют оболочки бактерий и циа нобионтов, панцири, раковины и скелеты животных, а также ткани, оболочки спор и пыльцы растений. В органическом породробразовании самую большую роль играют высшие растения. Их массовые скопления при определенных процессах захоронения приводят к возникновению горючих ископаемых (каустобиолитов), таких как торф, уголь, горючие сланцы, нефть и газ. Происхождение нефти и газа связано с глубоким разрушением первичного органического состава, вызванным как жизнедеятельностью бактерий и цианоби онтов, так и геологическими процессами. За счет жизнедеятель ности высших растений образуются смолы (янтарь).
Необходимо отметить и рельефообразующую роль ископае мых и современных организмов, создающих рифовые постройки различного типа: береговые и барьерные рифы, атоллы, биостромы, биогермы и др. Рифовые постройки имеют сложное строе ние. Они состоят из комплекса взаимозамещающих известковых пород: органогенных, обломочно-органогенных и хемогенных.
Рифовые постройки возвышаются в рельефе в виде гряд, хол мов и других поднятий. Их превышение над синхронными обра зованиями может быть довольно значительным. В образовании ископаемых и современных рифов принимают участие различ ные организмы (рис. 4). При восстановлении условий образования коралловых построек необходимо иметь в виду, что они могут быть и глубоководными. Иначе появляются ложные палеоэкологи ческие и тектонические реконструкции. Основное отличие глубо ководных от мелководных коралловых рифовых тел заключается
вотсутствии растений и волновой эрозии. В результате цементом
вглубоководных коралловых постройках служит известковый
15