

Физиология возбудимых тканей II
.pdfМинистерство здравоохранения Российской Федерации Санкт-Петербургский государственный медицинский университет имени академика И.П.Павлова
Кафедра нормальной физиологии
А.М.Коробкова, А.Ю.Юров
ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
Часть II
Пособие для студентов лечебного, стоматологического факультетов
и факультета спортивной медицины
Санкт-Петербург
2002
А в т о р ы : к.б.н. А.М.Коробкова, к.б.н. А.Ю.Юров.
Р е ц е н з е н т ы : проф. Р.В.Бабаханян,
проф. А.К.Косоуров.
Утверждено на заседании ЦМ К по химико-физиологическим дисциплинам. Протокол №3 от 10 января 2002 г.
В пособии рассматриваются механизмы синаптической передачи, работа скелетной, гладкой и сердечной мышц. Пособие содержит проверенные фундаментальные знания в объеме, соответствующем учебным планам СПбГМ У и медицинскому стандарту USM LE. Предназначено для студентов II курса лечебного, спортивного и стоматологического факультетов.
Издательство СПбГМУ, 2002
СИНАПТИЧЕСКАЯ ПЕРЕДАЧА ВОЗБУЖДЕНИЯ
Передача информации в нервной системе осуществляется в виде распространения возбуждения (ПД) по нервным волокнам. Проведение во збуждения по нервному волокну предусматривает его непрерывность. Однако нервная система представляет собой не морфологический синцитий, как предполагалось ранее, а представлена, как и любая ткань, отдельными клетками – нейронами. Это было доказано в конце ХIХ века работами итальянского врача и гистолога Л.Гольджи, применившего метод окрашивания срезов нервной ткани солями серебра. В подавляющем большинстве случаев мембраны нейронов не имеют контакта друг с другом. Следовательно, проведение ПД с одной клетки на другую по механизму проведения ПД в целостном волокне становится невозможным. В этом случае передача ПД осуществляется через синапс.
Основные положения синаптической передачи информации заключаются в следующем.
1.Синапс – это морфологическое образование, в котором электрическая активность одного нейрона изменяет активность другого нейрона (или иной клетки).
2.В некоторых случаях синаптическая передача может осуществляться электрически через прилегающие мембраны двух клеток.
3. В большинстве случаев синаптическая передача осуществляется с помощью химического вещества передающей клетки, которое называется медиатор. В таких синапсах электрический сигнал преобразуется в химический и химический вновь преобразуется в электрический.
4.Потенциал действия (ПД) в передающей информацию клетке высвобождает медиатор в синаптическую щель, которая разделяет две клет-ки
–передающую и принимающую сигнал.
5.На мембране воспринимающей сигнал клетки возникает изменение потенциала – либо деполяризация, либо гиперполяризация.
6.Если клетка деполяризуется и ее мембранный потенциал приближается в порогу возникновения потенциала действия, то синапс с таким механизмом изменения мембраного потенциала называют возбуждающим.
7.Если клетка гиперполяризуется и ее мембранный потенциал удаляется от порога возникновения потенциала действия, то такой синапс называют тормозным.
-4 -
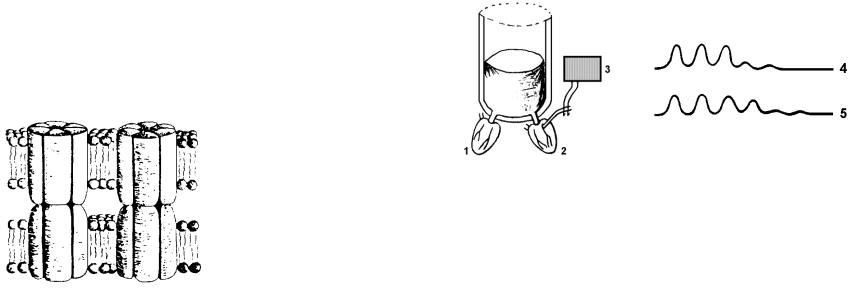
ЭЛЕКТРИЧЕСКИЕ СИНАПСЫ
Электрические синапсы встречаются в организме человека очень редко, только в сетчатке и в обонятельной луковице. Электрическая пер едача возбуждения совершается пассивным электротоническим распр остранением тока между мембранами двух клеток через щелевые соединения (рис.1). Мембраны двух клеток разделены щелью в 2 нм. Между ними создаются гидрофильные каналы-мостики из шести молекул интегральных белковконнексинов. Небольшие молекулы и ионы могут перемещаться через эти каналы из одной клетки в другую. Электрический ток, протекающий по мембране, через щелевой контакт свободно распространяется в любом направлении на мембрану другой клетки В электрических синапсах передача возбуждения осуществляется быстрее, чем в химических синапсах, и является надежной, если величина тока достаточно большая. Эти синапсы практически не утомляются, т.е. не теряют способности к выполнению своих функций. Но очень малое время последействия в электрических синапсах недостаточно для суммации, т.е. слабый сигнал, не способный возбудить соседнюю мембрану, не может быть многократно усилен сложением нескольких слабых сигналов и не передает информацию.
Рис.1. Щелевые контакты в электрических синапсах (Хухо, 1990).
Хотя в нервной системе человека этот тип передачи информации не имеет широкого распространения, электрические контакты широко распространены в организме человека там, где биологически целесообразно одновременное возбуждение большого массива клеток, например в гладкой мышечной ткани, в сердечной ткани.
- 5 -
ХИМИЧЕСКИЕ СИНАПСЫ
Наличие химической передачи возбуждения от нервной клетки было предположено в начале ХХ века и доказано в опытах О. Леви (1921). Результат эксперимента (рис.2) можно было объяснить только выделением возбужденным нервом какого-то биологически активного вещества, распространением этого вещества в растворе и воздействием не только на иннервируемое этим нервом сердце, но и на другое, которое омывается тем же раствором. Электронномикроскопические исследования в сочетании с данными гистохимии и электрофизиологии помогли в дальнейшем изучить структуру и химические свойства межклеточных контактов, в которых используются биологически активные молекулы.
А |
Б |
Рис.2. Опыты Леви, доказывающие наличие химических медиаторов в синапсах. А – схема опыта, Б – кривая сердечных сокращений при раздражении блу ждающего нерва: 1 – денервированное сердце лягушки, 2 – сердце лягушки с оставшейся веточкой блуждающего нерва, 3 – стимулятор, 4 – вагусная остановка раздражаемого сердца, 5 – остановка второго сердца.
Нервно-мышечный (мионевральный) синапс. Одним из наиболее изу-
ченных синапсов с химической передачей является возбуждающий синапс между мотонейроном спинного мозга и быстрым фазным волокном скелетной мышцы (рис.3).
Аксон мотонейрона широко ветвится и находится в углублении, обр а- зованном мембраной мышечного волокна. Передача импульса осуществляется от мембраны окончания аксона, которая называется пресинаптической мембраной, на мембрану мышечного волокна, которая называется
постсинаптической мембраной. Обе мембраны разделены синаптичес-
- 6 -

кой щелью. Сложное строение синапса придает ему механическую усто й- чивость и не позволяет мембранам смещаться относительно друг друга при сокращении мышцы.
А
Б |
В |
Рис.3. Строение нервно-мышечного синапса (Bullock et al., 1995).
В пресинаптической терминали (окончании аксона, который передает возбуждение) находятся синаптические пузырьки размером около 50 нм. Они содержат химический посредник (медиатор) – ацетилхолин (АХ), в каждом пузырьке примерно 5-10 тысяч молекул. Синтез АХ происходит в пресинаптической терминали из холина и ацетил-Ко-А с помощью фермента ацетилтрансферазы. В самой мембране выделяются активные зоны и зоны прикрепления. Пузырьки с медиатором сосредоточены возле актив-
- 7 -
ных зон. Мембрана активной зоны имеет потенциалзависимые Са++-каналы, которые открываются при деполяризации пресинаптической мембр аны.
Высвобождение АХ происходит путем экзоцитоза, когда пузырек связывается с зоной прикрепления, сливается с пресинаптической мембр аной и медиатор перемещается в синаптическую щель. Распространяющийся по аксону ПД деполяризует пресинаптическую мембрану. В активных зонах происходит открывание потенциалзависимых Са++-каналов. Са++ по градиенту концентрации входит в клетку и, по одной из версий, дефосфорилирует белок синапсин, связанный с пузырьками, содержащими медиатор. Блокирование экзоцитоза синапсином прекращается. Одновременно 200-300 пузырьков присоединяются к зоне прикрепления пресинаптической мембраны и в синаптическую щель выделяется огромное количество медиатора.
Синаптическая щель имеет размер около 60 нм. Она заполнена аморфной сетью из соединительной ткани, в которой находится фермент ацетил- холин-эcтераза (АХЭ), расщепляющий АХ. АХЭ синтезируется мышечной клеткой и с помощью коллагеноподобного "хвоста" прикрепляется к базальной мембране, или – сарколемме, отделяющей нервное окончание от мембраны мышечной клетки. Одна молекула этого фермента способна нейтрализовать до 10 молекул АХ за 1 мс.
Постсинаптическая мембрана имеет белки-рецепторы, состояние которых не зависит от величины потенциала на мембране, но зависит от связывания с медиатором. Такие рецепторы хемозависимы. Они активируются как АХ, высвобождающимся в синаптическую щель, так и алкалоидом табака никотином, поэтому называются никотин-чувствительными рецепторами.
Белок состоит из 5 субъединиц. Две из них являются собственно рецептором – местом распознавания и связывания АХ, три оставшиеся субъединицы представляют собой гидратированный канал, меняющий свою ко н- формацию, и в равной мере проницаемый как для К+, так и для Na+.
Когда рецептор соединяется с АХ и канал открывается, начинается движение ионов по электрохимическому градиенту. Na+ перемещается внутрь, а К+ – наружу. Токи для этих ионов рассчитываются по закону Ома:
Ik = gk(Em -Ek ), INa = gNa(Em –ENa ),
где gk, gNa – проводимость для К+ и Na+; Em – мембранный потенциал;
Ek, ENa – равновесные потенциалы для К+ и Na+. - 8 -

Поскольку gNa=gK, а электродвижущая сила (Em -ENa) для натрия больше
(так как равновесный потенциал для натрия +68 мВ, а для калия |
-90 мВ), |
входящий ток натрия преобладает и мембрана деполяризуется. Эта деполя-
ризация называется возбуждающий постсинаптический потенциал
(ВПСП). В нервно-мышечном синапсе ВПСП имеет собственное название –
потенциал концевой пластинки (ПКП). По мере деполяри-зации мембра-
ны электродвижущая сила для Na+ уменьшается, а для K+ увеличивается и в итоге возникает состояние равновесия, когда входящий и выходящие токи через мембрану равны и противоположны. Мембранный потенциал, при котором IK = INa и суммарный ток равен нулю, называется потенциал реверсии тока концевой пластинки, он равен примерно -15-20 мВ. ПКП не может быть меньше потенциала реверсии (как и любой градуальный потенциал мембраны). Постсинаптическая мембрана фазного скелетного м ы- шечного волокна деполяризуется до потенциала реверсии при однократном ПД аксона.
ПКП зависит от силы раздражения, в данном случае – от количества выделившегося медиатора. Одна молекула АХ открывает один канал и деполяризует мембрану на несколько мкВ. Один пузырек с 5-10 тыс. молекул деполяризует мембрану на 1 мВ в течение 1 мс. Подобный ответ регистрируется на постсинаптической мембране в покое, когда спонтанно выделяется медиатор из одного пузырька. Этот потенциал называется миниатюрный потенциал концевой пластинки (мПКП). мПКП регистрируются в по-
кое постоянно, это естественная не блокируемая утечка медиатора. Возможно, она служит для поддержания нормального состояния мышцы, так как денервация практически сразу приводит к атрофии м ышечного волокна.
На постсинаптической мембране нет потенциал-зависимых каналов, и ПД на ней невозможен.
Потенциалзависимые каналы находятся на соседних участках мембраны, куда ПКП распространяется электротонически от постсинаптической мембраны. Его величина достаточна для деполяризации этих участков до критического уровня и генерации в них ПД. Распространение ПД по мембране мышечного волокна аналогично проведению возбуждения по немиелинизированному нервному волокну.
После завершения синаптической передачи все измененные параметры в структурах синапса восстанавливаются. Са++ после высвобождения медиатора связывается с внутриклеточными органеллами в пресинаптичес-
- 9 -
кой терминали, аккумулируется в гладкой ЭПС, митохондриях, выводится из клетки работой Са++-АТФазы или Са++/Na+-обменом и последующим выведением Na+ работой Na+/K+-АТФазы с затратой энергии. Восстановление прежней по размерам поверхности пресинаптической мембраны происходит путем эндоцитоза с захватом холина из пресинапттической щели. Образовавшиеся пузырьки заполняются вновь синтезированным АХ.
Связь АХ и холинорецептора распадается и АХ разлагается ферментом АХЭ до ацетата и холина, который возвращается обратно в пресинаптическую терминаль и используется для нового синтеза АХ. Инактивация медиатора необходима для предотвращения длительной мышечной тетании (длительного сокращения, вызванного высокой частотой ПД в м ышечном волокне). Если инактивация АХ по каким-то причинам невозможна, рецепторы не теряют способность соединяться с медиатором, но теряют способность к изменению конформации и инактивируются. Это явление называется десенситизацией холинорецепторов. Механизм его не выяснен. Никотин и другие фармакологические препараты и токсины, которые могут открывать холинорецепторы в постсинаптической мембране мышечного волокна, десенситизации не вызывают, поэтому отравления этими веществами особо опасны.
Восстановление ионных градиентов Na+/K+ после трансмембранных токов осуществляется Na+/K+-АТФазой.
ОСОБЕННОСТИ ВОЗБУЖДЕНИЯ В ЦЕНТРАЛЬНОМ СИНАПСЕ
Центральный возбуждающий синапс. Синапсы между нейронами в центральной нервной системе называют центральными синапсами. Они образуются между различными структурами нейрона и могут разделяться на аксо-дендритные, аксо-соматические, аксо-аксональные (рис.4).
В центральной нервной системе есть как возбуждающие, так и тормозные синапсы. Порядок передачи ПД в возбуждающем синапсе принципиально мало отличается от передачи ПД в нервно-мышечном синапсе, но:
1.Возбуждающими медиаторами являются глутамат, аспартат, АХ, вещество Р и другие биологически активные вещества.
2.Рецептор редко сам является ионным каналом. Комплекс рецептормедиатор активирует сложные многоступенчатые метаболические про - цессы в клетке, изменение ионной проницаемости мембраны является последней ступенью в этих процессах.
-10 -
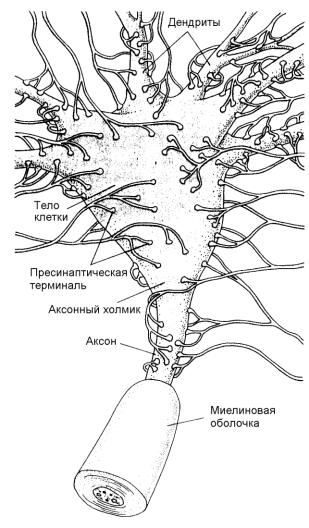
Рис.4. Центральные синапсы, образованные на одном нейроне (Sherwood,
1990).
- 11 -
3.ПД в случае синапса между аксоном и сомой возникает в аксонном
холмике, потому что аксонный холмик не содержит синапсов, имеет максимальную плотность расположения потенциалзависимых Na++-каналов, порог возбуждения мембраны аксонного холмика примерно в три раза ниже, чем у соматической мембраны.
4.После однократного стимулированного высвобождения медиатора амплитуда ВПСП недостаточна для генерации ПД в аксонном холмике. Для достижения критического потенциала необходимо суммирование ВПСП, последовательно развивающихся в одном синапсе (временная суммация) или одновременно в нескольких синапсах на одном нейроне (пространственная суммация). Поскольку процессы развития ВПСП длительные, суммация и возрастание амплитуды ВПСП являются обычным механизмом для центрального синапса.
Общее количество синапсов на одной нервной клетке велико, соотно - шение количества возбуждающих и тормозных синапсов различно. Одновременно в клетке активны десятки синапсов, т.е. трансмембранные токи ионов осуществляются через множество каналов и общий мембранный эффект – деполяризация или гиперполяризация – определяется суммированием всех ионных токов. Если все деполяризующие и гиперполяризующие ионные токи равны, то МП клетки, в том числе и аксоннного холмика, не меняется (рис.5).
5.Инактивация медиатора происходит после его диффузии в окружающем пространстве или в цитоплазматических структурах нейрона после активного поглощения медиатора обратно через пресинаптическую мембрану (re-uptake). Для некоторых медиаторов, в частности АХ, существует ферментативное расщепление в синаптической щели, как и в нер вно-мы- шечном синапсе.
Центральный тормозный синапс. Тормозные синапсы препятствуют возбуждению постсинаптической терминали. Они отличаются от во збуждающих синапсов тем, что:
1)медиаторами чаще всего являются глицин или ГАМК;
2)медиатор вызывает не деполяризацию, а гиперполяризацию постсинаптической мембраны, МП удаляется от порогового потенциала для генерации распространяющегося возбуждения и происходит торможение нейрона. Такой потенциал называется тормозным постсинаптическим потенциалом (ТПСП), а механизм торможения называется постсинапти-
ческим торможением.
-12 -

Рис.5. Суммация возбуждающих и тормозных постсинаптических потенциалов
(Sherwood, 1990).
Во время действия тормозного медиатора изменяется проводимость мембраны для ионов К+ и, если мембранный потенциал был меньше равновесного потенциала для К+, то усиливается выходящий ток К+, гиперполяризуя мембрану до -90 мВ. При гиперполяризации мембраны от небольших значений МПП до -80 мВ (равновесный потенциал для ионов хлора) возрастает также вход ионов хлора в клетку.
- 13 -
В аксо-аксональных контактах возможно гиперполяризационное или деполяризационное пресинаптическое торможение. Оно всегда ингибирует возбуждающий синапс, уменьшая количество Са++, входящего в пресинаптическую терминаль во время синаптической передачи, и выход во збуждающего медиатора (рис.6). Медиатором таких синапсов является ГАМК, но механизм ее действия зависит от типа рецепторов, которые присутствуют на пресинаптической терминали.
Рис.6. Пресинаптическое торможение (Sherwood, 1990).
- 14 -
А. Когда ГАМК связывается с рецепторами типа А, проводимость мембраны нейрона для С1– увеличивается:
1)если МПП меньше -80 мВ (равновесного потенциала для хлора), ионы Сl– входят в клетку. Мембрана гиперполяризуется до -80 мВ и порог возбуждения повышается;
2)если МПП -90 мВ, ионы С1– выходят из клетки, мембрана деполяризуется до -80 мВ и длительная деполяризация вызывает инактивацию Na+- каналов и уменьшение возбудимости.
ПД по возбуждающему волокну достигает пресинаптической терм ина-
ли, но его амплитуда снижена, потому что в первом случае входящий ток хлора через открытые хлорные каналы шунтирует входящий ток Na+, во втором случае большая часть Na+-каналов инактивирована. Поскольку амплитуда ПД падает, уменьшается количество открывающихся Са++-каналов и меньше ионов Са++ входит в пресинаптическую клетку. Выброс медиатора и амплитуда ВПСП понижаются.
Б. Если ГАМК связывается с рецепторами типа В, активируется G- протеин мембраны и уменьшение количества медиатора происходит двумя возможными путями:
1) G-белок усиливает выход К+ через открываемые им К+-каналы, порог раздражения повышается и/или амплитуда ПД уменьшается из-за нейтрализации входящего Na+ выходящим током К+;
2) G-белок прямо ингибирует Са++-каналы и уменьшает вход Са++ в пресинаптическую терминаль.
Наибольший тормозный эффект получается, если тормозный импульс приходит в аксо-аксональный синапс за несколько мс до прихода возбуждающего импульса.
При пресинаптическом ингибировании возбудимость и другие сво йства постсинаптической мембраны не изменяются. Аксо -аксональное соединение эффективно тормозит все приходящие к ней возбуждающие импульсы в пресинаптическом аксоне. Пресинаптическое ингбирование позволяет блокировать один возбуждающий синапс, не влияя на способность других синапсов возбуждать клетку. Это один из примеров взаимного во здействия синапсов друг на друга. Особенно часто пресинаптическое то рможение встречается у аксонов первичных чувствительных нейронов. Оно ограничивает приток избыточной информации в нервную систему.
Синаптическая пластичность. Работа синапса не является неизменной. При длительном ритмическом воздействии множества ПД на синапс наблюдается два эффекта.
- 15 -
1. Облегчение (сенситизация, посттетаническая потенциация),
характерное для начала стимуляции. При частом ритме ПД в пресинаптической терминали накапливается какое-то количество Са++, который увеличивает количество выделяющегося медиатора.
2. Депрессия, возникающая при длительной стимуляции вследствие истощения запасов медиатора в пресинаптической терминали. Это явление в некоторой степени компенсируется балансом между расходом и "поступлением + новым синтезом" медиатора, поэтому синапсы могут длительное время работать на сбалансированном уровне. Скорость синтеза, скорость транспорта медиатора в пресинаптическую терминаль, обратный захват медиатора из синаптической щели являются важнейшими факторами, регулирующими запас медиатора в синапсе.
Явления облегчения и депрессии в некоторых синапсах ЦНС являются
факторами синаптической пластичности. Облегчение участвует в "про-
торении" новых путей при формировании временных связей, выр аботке условных рефлексов, в других процессах памяти, а депр ессия является одним из механизмов адаптации – потери чувствительности к раздражителю при его длительном действии.
МЕДИАТОРЫ
Для работы химического синапса необходим медиатор. Медиаторы – это низкомолекулярные водорастворимые амины или аминокислоты и ро д- ственные им вещества (АХ, катехоламины – АДР, НАДР, дофамин, серотонин, аминокислоты – глицин, глутамат, ГАМК, таурин и пептиды). АХ и катехоламины синтезируются из циркулирующих в крови предшественников, аминокислоты и пептиды на раннем этапе синтеза связаны с глюкозой (табл.).
В последние годы много внимания уделено роли низкомолекулярных нейропептидов в нервной системе и в синаптической передаче. Это вещества гипоталамуса (вазопрессин, окситоцин, рилизинг-факторы: либерины и статины), других отделов ЦНС (энкефалины, эндорфины), половые гормоны, ангиотензин, простагландины и др. Часть из них была впервые о бнаружена даже не в ЦНС, а в желудочно-кишечном тракте (вещество Р, ВИП). Всего известно несколько десятков таких веществ. Они названы нейромодуляторами, поскольку обладают некоторыми свойствами м едиаторов. Нейромодуляторы не оказывают прямого медиаторного эффекта, но также, как и медиаторы, они должны находиться в синапсе, изменять его
- 16 -
- 17 - |
- 18 - |
работу, иметь свои органы-мишени и свои механизмы инактивации, вызывать эффект при внесении их в синапс в физиологической дозе.
Идентификация, химический анализ и доказательства физиологический роли медиатора – труднейшая задача. Критерии, согласно которым вещество является медиатором, следующие.
1.Анатомический – присутствие данного вещества в подходящих количествах в пресинаптических окончаниях.
2.Биохимический – присутствие и активность ферментов, которые синтезируют данное вещество в пресинаптическом нейроне и его отростках
иудаляют или инактивируют это вещество в синапсе.
3.Физиологический – физиологическая стимуляция заставляет пресинаптическое окончание выделять данное вещество, а его ионофоретич еская инъекция в естественных концентрациях в область синапса имитирует естественную реакцию активного синапса.
4.Фармакологический – фармакологические препараты, которые влияют на различные ферментативные или биофизические этапы, оказывают ожидаемое действие на синтез, накопление, высвобождение, активность, инактивацию и обратный захват данного вещества.
Наиболее полно этим критериям удовлетворяет АХ в нервно -мышеч- ном синапсе. Другие вещества, признаваемые медиаторами (см. табл.), бо - лее или менее приближаются к этим критериям.
Принцип Дейла. Нервная система использует разные вещества в качестве медиаторов в разных синапсах. Однако во множестве синапсов, об - разованных только одним нейроном, медиатор, как правило, один и тот же. Метаболическое единство нейрона требует, чтобы один нейрон мог транспортировать и выделять синтезированный им медиатор в окончаниях всех
своих отростков: как ветвящегося аксона, так и дендритов. Согласно у т- верждению Дейла, если некоторое вещество в одном синапсе будет идентифицировано как медиатор, то можно будет заключить, что оно же будет медиатором и во всех других синапсах, образованных этим нейроном.
Принцип Дейла не утверждает, что в одной клетке может синтезир о- ваться только один медиатор. Достоверно доказано, что в нейронах улитки аплизии есть четыре медиатора. Все эти медиаторы транспортируются во все отростки нейрона. В нейронах человека также присутствуют несколько медиаторов.
Принцип Дейла вовсе не означает, что результаты действия всех синапсов одного нейрона идентичны во всех постсинаптических клетках. Ха-
- 19 -
рактер взаимоотношений между медиаторами и вызываемыми ими эффектами наиболее полно сформулирован Э.Кэнделом в 1976 году.
1.Знак синаптического действия определяется не только медиатором, но и свойствами рецепторов на постсинаптической мембране
(АХ возбуждает клетки желудочно-кишечного тракта и усиливает их сокращение и секрецию, но тормозит работу сердца, гиперполяризуя клеткиводители ритма).
2.Рецепторы на клетках, являющихся постсинаптическими по отношению к одному пресинаптическому нейрону, могут фармаколо-
гически различаться и могут контролировать разные ионные каналы
( 1-адренорецепторы сердца при связывании с медиатором адреналином увеличивают вход Са++ в кардиомиоциты и сила сокращений сердца возрастает, но 2-адренорецепторы гладкой мускулатуры бронхов при связывании
садреналином усиливают только перемещение Са++ в гладкую ЭПС и митохондрии и способствуют расслаблению бронхов).
3.Одна постсинаптическая клетка может иметь более одного типа рецепторов для данного медиатора и каждый из этих рецепторов
может контролировать отличный от других механизм ионной прово-
димости ( 1-адренорецепторы кровеносных сосудов активируют вход Са++ и сокращение мышечных клеток стенки сосуда, 2-адренорецепторы в этих же клетках, как и в гладких миоцитах бронхов, усиливают вход Са++ в гладкую ЭПС и вызывают расслабление мышц).
Резюме
Общие свойства химической передачи возбуждения:
1)односторонний характер проведения в синапсе обеспечивает пер е- дачу информации от рецептора к анализатору, от анализатора к эффектору;
2)возможно как возбуждение, так и торможение работы эффектора;
3)использование в различных сочетаниях небольшого количества м е- диаторов со столь же небольшим количеством рецепторов позволяет вызывать разнообразные ответы в эффекторах;
4)возможно образование функциональных систем синапсов (суммация, пресинаптическое ингибирование);
5)изменение частоты возбуждающих потенциалов действия в аксонном холмике нейрона в зависимости от суммарной величины ПСП дает возможность для кодирования сенсорной информации и интегративной деятельности нейрона;
-20 -