

34. Онто-филогенез головного мозга.
Головной мозг — encephalon — головная часть центрального отдела нервной системы, расположенная в полости черепа. Развитие его, как и спинного мозга, произошло путем замыкания краев нервной борозды с образованием нервной трубки. Высокая концентрация нервной ткани вокруг пяти пузырей на переднем конце тела привела к развитию разнообразных по строению и функции структур.
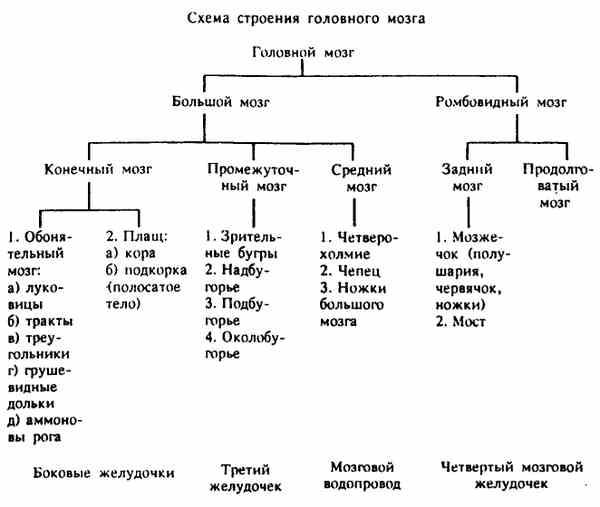
Схема 1. Строения головного мозга
Так, конечный мозг наиболее богат сложными ассоциативными структурами, тесно связан с органами чувств наиболее древнего нервно-эпителиального типа.
Головной мозг с дорсальной поверхности разделяется поперечной щелью — fissura trausversa cerebri на большой и ромбовидный мозг.
Большой мозг — cerebrum. Состоит из конечного, промежуточного и среднего мозга, имеет два полушария, представляющих конечный мозг. Правое и левое полушария большого мозга — hemispheria cerebri dextrum et sinistrum дорсально разграничиваются глубокой продольной щелью — fissura longitudinalis cerebri. Полушария прикрывают с дорсальной поверхности промежуточный и средний мозг. С базальной стороны можно видеть части промежуточного мозга — гипофиз и перекрест зрительных нервов, а также среднего мозга — ножки большого мозга.
Ромбовидный мозг — rhombencephalon. Состоит из заднего мозга, в который входят мозжечок и мозговой мост, и продолговатого мозга. Мозжечок — cerebellum располагается дорсально от продолговатого мозга и позади от полушарий большого мозга. На переднем конце продолговатого мозга с вентральной поверхности лежит мозговой мост — pons. Продолговатый мозг — medulla oblongata, s. myelencephalon непосредственно продолжается в спинной мозг.
У рогатого скота большой мозг сравнительно короткий, широкий и высокий; полушария спереди сужены, сзади сильно расширены, что придает мозгу грушевидную форму.
У лошадей большой мозг сравнительно длинный, более сжат с боков и ниже, чем у жвачных. Извилин больше, чем у рогатого скота.
У свиней сильно развиты латеральные обонятельные тракты, а дугообразные борозды не такие четкие, как у собаки.
У собак общая форма головного мозга зависит от очертаний черепа: она то более грушевидная, то более круглая. Для мозга собаки типичны три дугообразные борозды на плаще.
Масса головного мозга новорожденных отличается от массы мозга взрослых, у разных животных она неодинакова.
Морские свинки, лошади, крупный рогатый скот, овцы рождаются «зрелыми», появляются на свет с массой головного мозга, составляющей от массы мозга взрослых особей 40—60%, тогда как собаки, кошки, кролики, крысы и мыши — животные, неспособные сразу после рождения к активному существованию (к самостоятельному передвижению, питанию), имеют относительно малый по размерам мозг—13—17% от массы мозга взрослых животных. Самым не-зрелым (12%) в этом ряду оказывается мозг ребенка.
Развитие головного мозга. На ранних стадиях онто-и филогенеза головной мозг представляет расширенный конец мозговой трубки, который называется первичным, или прехордальным, мозговым пузырем, так как он лежит впереди хорды. Развитие его функционально связано с органом обоняния, и его обонятельная функция сохраняется у всех животных, включая млекопитающих. Несколько позже позади прехордального мозгового пузыря на переднем конце мозговой трубки появляется вторичный мозговой пузырь (его называют также эпихордальным мозговым пузырем, так как он лежит дорсально от хорды). Развитие вторичного мозгового пузыря обусловлено:
а)функцией жаберного аппарата и органов боковой линии водных животных (соответственно органу равновесия у наземных животных);
б)дифференцировкой внутренних органов, которая, в свою очередь, была вызвана повышением интенсивности обмена веществ и усилением подвижности животных.
в)возникновением первичного ассоциативного и коммиссурального центра. С усовершенствованием органа зрения от эпихордального мозгового пузыря обособляется средний мозговой пузырь — mesencephalon. На этой трехпузырной стадии прехордальный мозговой пузырь именуют передним мозгом — prosencephalon, а эпихордальный мозговой пузырь — задним, или ромбовидным, мозгом — rhombencephalon. С дорсальной поверхности все три отдела мозга довольно резко отграничиваются друг от друга поперечными коммиссурами, или спайками, из нервных волокон как впереди, так и позади среднего мозга. В дальнейшем возникает пятипузырная стадия мозга. Из прехордального мозга образуется конечный мозг — telencephalon в виде парного мозгового пузыря и промежуточный мозг — diencephalon.
Наивысшей ступени развития, особенно в области, плаща, конечный мозг достигает у млекопитающих, в частности у человека. Промежуточный мозг у низших животных развит слабо, но затем с повышением организации животных он становится высшим подкорковым чувствительным центром, связанным со всеми рецепторами анализаторов и через спинной мозг со всеми органами тела, а также с корой большого мозга по мере ее разрастания. Из него же происходят глазные пузыри. Почти одновременно с делением прехордального мозга дифференцируется и ромбовидный мозг на задний мозг — metencephalon и продолговатый мозг — myelencephalon. Задний мозг первоначально представлен одним лишь мозжечком, который является подкорковым центром корреляции мышечных движений для сохранения равновесия. Лишь у млекопитающих к мозжечку вследствие диффе-ренцировки коры большого мозга добавляется мозговой мост. Продолговатый мозг — непосредственное производное эпюсордального мозгового пузыря, функции которого он и сохраняет. Полости первичных мозговых пузырей в развитом мозге становятся мозговыми желудочками. Из полости прехордального мозга в конечном мозге возникают парные боковые желудочки, а в промежуточном мозге — третий мозговой желудочек. Все три желудочка соединяются межжелудочковым отверстием (монроево). Желудочек среднего мозгового пузыря вследствие разрастания стенок последнего превращается в мозговой водопровод. Полость же эпихордального мозгового пузыря становится четвертым мозговым желудочком. Он сообщается и с центральным спинномозговым каналом, и с мозговым водопроводом. Развитие конечного мозга. Из передней и вентральной стенок конечного мозга происходят обонятельный мозг — обонятельные доли и полосатые тела, а из дорсальной его стенки — плащ. Развитие обонятельных долей мозга обусловливается наличием обонятельного анализатора, который у водных животных играет исключительную роль при ориентации во внешней среде. Отростки чувствительных обонятельных клеток заканчиваются в обонятельных долях — выростах стенки конечного мозга. Дистальные отделы обонятельных долей формируют первичные обонятельные центры — обонятельные луковицы, а проксимальные отделы — древнейшую обонятельную кору из серого мозгового вещества в виде обонятельных извилин и обонятельных треугольников; они возникают как вторичные обонятельные центры.
В вентральной же стенке конечного мозга дорсально от закладки очень рано появляется закладка крупноклеточного базального ганглия. Это первичный и притом высший двигательный центр. Назальный ганглий сохраняется и у млекопитающих животных в виде бледного ядра. Позднее у наземных животных разрастаются дополнительные мелкоклеточные ядра, образующие скорлупу (начиная с рептилий), а у млекопитающих еще хвостатое ядро. Оба объединяются под названием «полосатое тело». С появлением вторичного плаща полосатое тело пронизывается внутренней и наружной капсулами из проводящих путей, идущих в кору плаща и обратно. В эволюции плаща наблюдаются две формации различной функции и строения, не считая мембранообразного примитивного плаща, характерного для водных животных и построенного только из эпендимы. У наземных животных разрастание вторичного плаща вызывается внедрением в него новых нервных волокон (проекционных) из промежуточного мозга, являющихся проводниками различных анализаторов — кожного, зрительного, слухового, мышечного. Кора вторичного плаща в ряду животных чрезвычайно усложняется в строении, резко дифференцируются ее функции и структура, увеличиваются размеры. У крупных млекопитающих животных плащ обычно испещрен извилинами и бороздами. Извилины в ряду животных располагаются неодинаково. В одних случаях (у хищных, копытных) они в основном идут дугами вокруг поперечной сильвиевой борозды; у приматов и человека извилины образуют две системы — ложную и теменную. Обе системы разделены сильвиевой бороздой. У третьей группы животных сильвиева борозда отсутствует, а борозды идут в передней части мозга продольно, а в задней — поперечно. В силу сказанного абсолютная гомологизация извилин между разными отрядами животных крайне затруднительна, а в отдельных случаях невозможна. У мелких животных извилины вообще отсутствуют (животные с гладким головным мозгом). Наибольшее количество извилин насчитывается у слонов и китов, у мелких приматов извилины отсутствуют. В онтогенезе извилины появляются также не сразу, а в определенной последовательности.
Филогенетически все поля дифференцируются из примитивных четырех областей мозга низших животных (сумчатых и насекомоядных). Эти области представляют собой мозговые отделы анализаторов: передние доли коры являются двигательным анализатором, затылочные доли — зрительным и промежуточная между ними доля — кожным анализатором. У высших животных возникают новые ассоциационные области: лобная доля, а затем и височно-теменная, с наиболее крупным объемом у человека. Так, лобные доли у кролика составляют 2%, у кошки 3, у собаки 7, у обезьян 8—16, у человека 29% общей массы головного мозга.
Абсолютная масса головного мозга колеблется в широких пределах, а относительная его масса обратнопропорциональна массе животного.
Абсолютная масса мозга у китов 4600 - 7000 г, а относительная масса 1/10 000 - 1/14 000; у слонов соответственно 4300 - 5400 г и 1/375 - 1/560; у лошади 372 - 570 г и 1/480 - 1/1000; у КРС 410 - 550 г и 1/600 - 1/770; у свиней 96 - 145 г и 1/1200 - 1/1900; у собак 46 - 138 г и 1/30 - 1/400; у темной цепкой обезьяны 126 г и 1/15; у человека 1350 - 1450 г и 1/35 - 1/45. У молодых животных относительная масса мозга значительно выше, чем у взрослых: у 5-недельного львенка 1/18, а у взрослого льва 1/546; у новорожденного ребенка 1/8, а у взрослого человека до 1/35.
Процентное отношение серого мозгового вещества к белому у мелких животных выше, чем у крупных: у лошади 52,1 и 47,6%; у овцы 54,9 и 45,1; у собаки 61,1 и 38,9; у хряка 80,7 и 19,3; у человека 40 и 60%.