

Основные различия между прокариотами и эукариотами:
|
Характеристика |
Прокариоты |
Эукариоты |
|
Размеры клеток |
Диаметр в среднем составляет 0,5-5 мкм |
Диаметр обычно до 40 мкм; объем клетки, как правило, в 1000-10000 раз больше, чем у прокариот |
|
Форма |
Одноклеточные или нитчатые |
Одноклеточные, нитчатые или истинно многоклеточные |
|
Генетический материал |
Кольцевая ДНК находится в цитоплазме и ничем не защищена. Нет истинного ядра или хромосом. Нет ядрышка |
Линейные молекулы ДНК связаны с белками и РНК и образуют хромосомы внутри ядра. Внутри ядра находится ядрышко |
|
Синтез белка |
70S-рибосомы и мельче. Эндоплазматического ретикулума нет. (Синтез белка характеризуется и многими другими особенностями, в том числе и чувствительностью к антибиотикам; например, развитие прокариот ингибируется стрептомицином.) |
80S-рибосомы (крупнее). Рибосомы могут быть прикреплены к эндоплазматическому ретикулуму |
|
Органеллы |
Органелл мало. Ни одна из них не имеет оболочки (двойной мембраны). Внутренние мембраны встречаются редко; если они есть, то на них обычно протекают процессы дыхания или фотосинтеза |
Органелл много. Некоторые органеллы окружены двойной мембраной, например ядро, митохондрии, хлоропласты. Большое число органелл ограничено одинарной мембраной, например аппарат Гольджи, лизосомы, вакуоли, микротельца эндоплазматический ретикулум и т.д. |
|
Клеточные стенки |
Жесткие, содержат полисахариды и аминокислоты. Основной упрочняющий компонент - муреин |
У зеленых растений и грибов клеточные стенки жесткие и содержат полисахариды. Основной упрочняющий компонент клеточной стенки растений - целлюлоза, у грибов - хитин |
|
Жгутики |
Простые, микротрубочки. Находятся вне клетки (не окружены плазматической мембраной). Диаметр 20 нм |
Сложные, с расположением микротрубочек типа 9+2. Располагаются внутри клетки (окружены плазматической мембраной). Диаметр 200 нм |
|
Дыхание |
У бактерий происходит в мезосомах; у сине-зеленых водорослей - в цитоплазматических мембранах |
Аэробное дыхание происходит в митохондриях |
|
Фотосинтез |
Хлоропластов нет. Происходит в мембранах, не имеющих специфической упаковки |
В хлоропластах, содержащих специальные мембраны, которые обычно уложены в ламеллы или граны |
|
Фиксация азота |
Некоторые обладают такой особенностью |
Ни один организм не способен к фиксации азота |
Эукариотические клетки в среднем намного крупнее прокариотических, разница в объёме достигает тысяч раз. Клетки эукариот включают около десятка видов различных структур, известных как органоиды (или органеллы, что, правда, несколько искажает первоначальное значение этого термина), из которых многие отделены от цитоплазмы одной или несколькими мембранами (в прокариотических клетках внутренние органоиды, окруженные мембраной, встречаются редко). Ядро — это часть клетки, окружённая у эукариот двойной мембраной (двумя элементарными мембранами) и содержащая генетический материал: молекулы ДНК, «упакованные» в хромосомы. Ядро обычно одно, но бывают и многоядерные клетки.
Для клеток прокариот характерно отсутствие ядерной оболочки, ДНК упакована без участия гистонов. Тип питания осмотрофный.
Генетический материал прокариот представлен одной молекулой ДНК, замкнутой в кольцо, имеется только один репликон. В клетках отсутствуют органоиды, имеющие мембранное строение. В геноме могут присутствовать мобильные генетические элементы, а у некоторых прокариот (например, вольбахия) их содержится необычно много. Изучение бактерий привело к открытию горизонтального переноса генов, который был описан в Японии в 1959 г. Это процесс широко распространен среди прокариот, а также у некоторых эукариот. Открытие горизонтального переноса генов у прокариот заставило по другому взглянуть на эволюцию жизни. Ранее эволюционная теория базировалась на том, что виды не могут обмениваться наследственной информацией. Прокариоты могут обмениваться генами между собой непосредственно (конъюгация, трансформация) а также с помощью вирусов - бактериофагов (трансдукция).
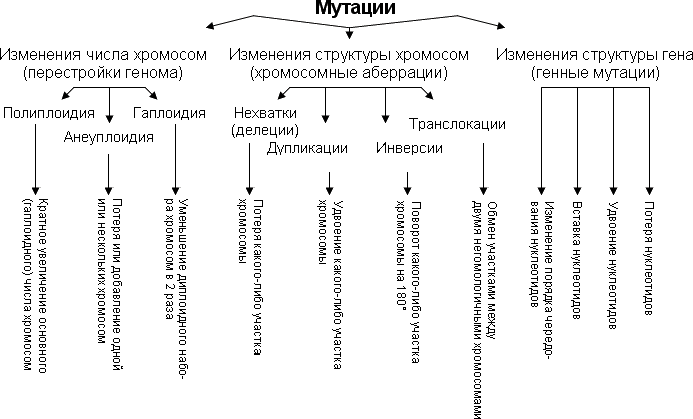
16.Классификация мутаций
Геномные мутации. В результате мутаций в ядре зиготы изменяется видовое число хромосом. Кариотип особи изучается на метафазных пластинках.Геномные мутации могут касаться всех хромосом (полиплоидия) или отдельных хромосом (анеуплоидия). В последнем случае может добавляться отдельная хромосома (трисомия) или вместо пары будет представлена одна хромосома (моногамия). Эти мутации редко оказываются жизнеспособными, чаще они приводят летальному исходу еще в процессе эмбриогенеза (спонтанные аборты), либо к рождению ребенка с нарушениями умственного и физического развития (врожденными пороками развития). Таковы синдромы анеуплоидии в виде моно- и трисомий по аутосомным и половым хромосомам. В частности, известный синдром Дауна обусловлен трисомией по 21-й паре хромосом. Синдром Дауна связан с нарушением ряда признаков - искаженные физические способности, умственная отсталость, выраженная от легкой дебильности до тяжелых форм идиотии. Частота данного заболевания в поколении 1 на 500-700 новорожденных.
Структурные мутации. В этом случае в результате мутации изменяется структура хромосомы. Основными видами структурных мутаций хромосом являются разрывные и обменные аберрации. К разрывным аберрациям относятся разного рода фрагменты (разделение хромосомы на части), к обменным аберрациям относятся случаи, когда имеются два разрыва и хромосома на их основе преобразуется, - транслокации (перенос участка хромосомы в другую хромосому или внутри хромосомы), инверсии (поворот участка внутри хромосомы на 180°), внутренние делеции (потеря внутренних участков хромосом), кольца (замыкание в кольцо отделяющегося внутреннего участка). Структурные изменения могут образовываться перемещающимися элементами в виде участков ДНК, мобильных по своему положению в организации генома. Как правило, структурные мутации хромосом приводят к множественным дефектам развития. Так, при делеции короткого плеча 5-й хромосомы (нехватке концевого фрагмента) наблюдается заболевание, названное синдромом "кошачьего крика". Помимо болезненного крика ребенка, напоминающего кошачье мяуканье, обусловленного аномалиями развития гортани, для больных детей характерны тяжелая умственная отсталость, задержка роста и другие симптомы. Ряд транслокаций и инверсий передаются потомкам.
Генные мутации. При повреждении или нарушениях в порядке или замене нуклеотидов, появлении внутренней дупликации или делеции в Молекуле ДНК возникают генные (точковые) мутации. Эти изменения отдельных генов часто приводят к тяжелым дегенеративным заболеваниям, в частности, многочисленным болезням обмена веществ через нарушения синтеза белков, ферментов. Примером может служить мутация, приводящая к появлению серповидноклеточной анемии — наследственного заболевания, как правило, приводящего детей и подростков к смерти. В этом случае в эритроцитах вместо нормального гемоглобина A содержится аномальный гемоглобин S. Аномалию вызывает мутация
Другой пример генной мутации - наследственное заболевание фенилкетонурия. Заболевание характеризуется резко выраженной умственной отсталостью, развивающейся вследствие нарушения нормальных биохимических процессов в мозге из-за накопления в организме фенилаланина. В отличие от нормальных детей у детей, больных фенилкетонурией, фенилаланин образуется при распаде тканевых белков и при переваривании белковой пищи. Мутация в гене, кодирующем фермент фенилала-нин-4-гидроксилазу, приводит к блокированию реакции превращения фенилаланина в тирозин.
Что касается вредных доминантных мутаций, то каждый из ее носителей специфически страдает от нарушений жизнеспособности. Таким образом, отрицательные доминантные гены прямо входят в состав генетического груза данного поколения из-за мутаций, которые возникают в зародышевых клетках родителей.
Механизмы возникновения:
Мутации по типу замены азотистых оснований. Эти мутации происходят в силу ряда конкретных причин. Одной из них может быть возникающее случайно или под влиянием конкретных химических агентов изменение структуры основания, уже включенного в спираль ДНК. Если такая измененная форма основания остается не замеченной ферментами репарации, то при ближайшем цикле репликации она может присоединять к себе другой нуклеотид. Примером может служить дезаминирование цитозина, превращающегося в урацил самопроизвольно или под влиянием азотистой кислоты .Образующийся при этом урацил, не замеченный ферментом ДНК-гликозилазой, при репликации соединяется с аденином, который впоследствии присоединяет тимидиловый нуклеотид. В результате пара Ц—Г замещается в ДНК парой Т—А. Дезаминирование метилированного цито зина превращает его в тимин Тимидиловый нуклеотид, являясь естественным компонентом ДНК, не обнаруживается ферментами репарации как изменение и при следующей репликации присоединяет адениловый нуклеотид. В результате вместо пары Ц—Г в молекуле ДНК также появляется пара Т—А.
Мутации со сдвигом рамки считывания. Этот тип мутаций составляет значительную долю спонтанных мутаций. Они происходят вследствие выпадения или вставки в нуклеотидную последовательность ДНК одной или нескольких пар комплементарных нуклеотидов. Большая часть изученных мутаций, вызывающих сдвиг рамки, обнаружена в последовательностях, состоящих из одинаковых нуклеотидов. Изменению числа нуклеотидных пар в цепи ДНК способствуют воздействия на генетический материал некоторых химических веществ, например акридиновых соединений.
Мутации по типу инверсии нуклеотидных последовательностей в гене. Данный тип мутаций происходит вследствие поворота участка ДНК на 180°. Обычно этому предшествует образование молекулой ДНК петли, в пределах которой репликация идет в направлении, обратном правильному. В пределах инвертированного участка нарушается считывание информации, в результате изменяется аминокислотная последовательность белка.
Эволюционное значение:
Мутации являются материалом для естественного отбора. Отбор не приводит к возникновению новых признаков, благодаря естественному отбору виды не возникают, а исчезают. Единственным источником видообразования есть мутационная изменчивость. Мутации происходят во всех возможных направлениях. Способность к мутациям наступает периодически.
17. Репарация — особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, повреждённой при нормальном биосинтезе ДНК в клетке или в результате воздействия физическими или химическими агентами.
Типы: Прямая репарация
Прямая репарация наиболее простой путь устранения повреждений в ДНК, в котором обычно задействованы специфические ферменты, способные быстро (как правило, в одну стадию) устранять соответствующее повреждение, восстанавливая исходную структуру нуклеотидов. Так действует, например, O6-метилгуанин-ДНК-метилтрансфераза, которая снимает метильную группу с азотистого основания на один из собственных остатков цистеина.
Эксцизионная репарация
Эксцизионная репарация (англ. excision — вырезание) включает удаление повреждённых азотистых оснований из ДНК и последующее восстановление нормальной структуры молекулы.
Пострепликативная репарация
Tип репарации, имеющей место в тех случаях, когда процесс эксцизионной репарации недостаточен для полного исправления повреждения: после репликации с образованием ДНК, содержащей поврежденные участки, образуются одноцепочечные бреши, заполняемые в процессе гомологичной рекомбинации при помощи белка RecA.
Пострепликативная репарация была открыта в клетках E.Coli, не способных выщеплять тиминовые димеры. Это единственный тип репарации, не имеющий этапа узнавания повреждения.