

Федеральное государственное автономное
образовательное учреждение высшего образования
«СИБИРСКИЙ ФЕДЕРАЛЬНЫЙ УНИВЕРСИТЕТ»
Институт фундаментальной биологии и биотехнологии
Базовая кафедра биотехнологии
Реферат
Тема: Классификация мобильных элементов, структура и способы перемещения
Красноярск 2018 г.
Содержание
СОДЕРЖАНИЕ 1
ВВЕДЕНИЕ 2
Глава I 3
Общее понятие МГЭ. История открытия 3
Эволюционное значение 4
Структура МГЭ 7
Способы перемещения 12
Введение
С момента возникновения хромосомной теории наследственности до конца 70-х годов представление о том, что каждый ген имеет определенное место на хромосоме и не способен произвольно менять его, казалось незыблемым. Единственным известным способом перемещения генов друг относительно друга были хромосомные мутации — транслокации и инверсии.
Другое очень распространенное и обоснованное представление гласит о том, что в геноме данного вида организмов содержится вполне определенное количество копий какого-либо конкретного гена. Изменение числа копий может также происходить в результате хромосомных мутаций — дупликаций и делений. В 40—50-х годах XX в. американская исследовательница Б. Мак-Клинток генетическими методами показала, что в хромосомах кукурузы предположительно существуют генетические элементы, способные перемещаться в геноме — исчезать с прежних мест и появляться в новых.
Спустя четверть века американские и советские генетики независимо методами молекулярной биологии и генной инженерии доказали существование генетических элементов, способных к перемещению.
Глава I
Общее понятие МГЭ. История открытия
Мобильные генетические элементы — это мобильные последовательности ДНК, найденные в геномах всех организмов. Эти элементы генома, имеют способность перемещаться из сайта в сайт, либо в процессе прямого вырезания вставки ДНК (по крайней мере, в прокариотах), либо путем транскрипции элемента, обратной транскрипции, образовавшейся РНК с образованием ДНК-копии или же ее внедрением в другое место генома. Во многих геномах они находятся в изобилии: например, они составляют до 50 % человеческой ДНК. Они часто вызывают мутации, либо вставляясь в другой ген и разрушая, или вызывая перестройки ДНК, такие как делеции, дупликации и инверсии. Когда мобильный генетический элемент, встраивается внутрь какого-то обычного гена, он нарушает его работу, вызывая мутацию исходного гена. Когда МГЭ меняет свою локализацию, нормальная работа гена возобновляется, мутация исчезает. Исходя из этого, была сформирована гипотеза, что «прыгающие гены» возникли из ДНК вирусов. Как известно, многие вирусы могут встраиваться в ДНК клетки хозяина, а потом покидать ее.
Открытие МГЭ принадлежит американскому генетику Барбаре Мак-Клинток, которая исследовала вариации окраски зерна и листьев кукурузы, и в 1948 году путём цитологических и генетических исследований пришла к выводу, что мобильные участки ДНК, Ac/Ds-элементы, приводят к соматическому мозаицизму растений. Она была первой, кто доказал, что геном эукариот не статичен, а содержит участки, которые могут передвигаться. В 1983 году за эту работу Барбара Макклинток получила Нобелевскую премию.
Хотя транспозоны были открыты в 1940-х годах, только через полвека стало понятно, насколько масштабным является их вклад в геном организмов. Так, получение первой нуклеотидной последовательности (секвенирование) генома человека показало, что мобильных элементов в последовательности ДНК не менее 50 %. Точную оценку получить трудно, поскольку некоторые транспозонные участки со временем настолько изменились, что их нельзя уверенно идентифицировать.
Поскольку транспозоны потенциально способны вызывать вредные мутации и поломки хроматина, с начала открытия мобильных элементов считалось, что их действие сводится к геномному паразитизму. Но в начале XXI столетия появляется всё больше данных о возможных благоприятных эффектах транспозонов для организмов, об эволюционном влиянии ретротранспозонов на геном плацентарных млекопитающих.
У человека транспозоны были обнаружены в 1991, когда Фрэнсис Коллинз и его коллеги обнаружили 31-летнего человека с нейрофиброматозом, вызванным перемещением последовательности Alu. Нейрофиброматоз — болезнь, которая вызывает многочисленные опухоли кожи и нервов. В настоящее время установлено, что от 45 до 50 % (по данным разных авторов) человеческого генома состоят из последовательностей, происходящих от мобильных элементов, хотя большинство этих элементов является бездействующими и не способны к перемещению. Из них, около 2 % — это ДНК транспозоны и приблизительно 42 % — ретротраспозоны.
Эволюционное значение
Эволюционное значение мобильных генетических элементов неизвестно, но были предложены три гипотезы, объясняющих их происхождение.
- Гипотезы «клеточной функции» предполагает, что мобильные элементы обеспечивают какую-то важную функцию клетки.
- Гипотеза «генетической изменчивости» предполагает, что мобильные элементы, вызывая мутации, обеспечивают эволюционную гибкость видов.
- Гипотеза «эгоистичной ДНК» предполагает, что мобильные элементы не приносят какую-либо пользу клетки, но они широко распространены из-за того, что они могут копироваться и распространяться.
Также установлена способность подвижных генетических элементов к точному вырезанию и удалению их из хромосом. Перемещение таких нуклеотидных последовательностей в пределах генома может влиять на регуляцию экспрессии генов, которые прилежат к месту встраивания этих элементов. В результате таких перемещений могут активироваться ранее не активные гены, и наоборот.
Некоторые этапы эволюционирования организмов были вызваны активностью мобильных элементов генома. Уже первая нуклеотидная последовательность генома человека доказала, что многие гены были производными транспозонов. Мобильные элементы генома могут влиять на организацию генома путём рекомбинации генетических последовательностей и входя в состав таких фундаментальных структурных элементов хроматина, как центромеры и теломеры. Мобильные элементы могут влиять на соседние гены, меняя узоры (паттерны) сплайсинга и полиаденилирования или выполняя функции энхансеров или промоторов. Транспозоны могут влиять на структуру и функции генов путём выключения и изменения функций, изменения структуры генов, мобилизации и реорганизации фрагментов генов и изменения эпигенетического контроля генов.
Репликация транспозонов может вызвать некоторые заболевания, но, несмотря на это, в процессе эволюции транспозоны не были удалены и остались в ДНК-последовательностях почти всех организмов, или в виде целых копий, которые имели возможность передвигаться по ДНК, или в укороченном виде, потеряв способность к передвижению. Но укороченные копии также могут принимать участие в таких процессах, как пост-транскрипционная регуляция генов, рекомбинация и т. п. Также важным моментом в потенциальной способности транспозонов влиять на темпы эволюции является то, что их регуляция зависит от эпигенетических факторов. Это приводит к возможности транспозонов реагировать на изменения окружающей среды и вызывать генетическую нестабильность. У растений мобильные генетические элементы очень чувствительны к различным типам стресса, на их активность могут влиять многочисленные абиотические и биотические факторы, среди которых солёность, ранения, холод, тепло, бактериальные и вирусные инфекции. По состоянию на 2012 год задокументировано 96 различных заболеваний человека, причиной которых является de novo внедрение мобильных генетических элементов.
Ещё одним возможным механизмом эволюции геномов организмов является горизонтальный перенос генов — процесс передачи генов между организмами, которые не находятся в отношениях «предки-потомки». Есть сведения о том, что взаимодействия паразитических организмов и животных-хозяев могут повлечь горизонтальный перенос генов с помощью транспозонов, который состоялся между позвоночными и беспозвоночными организмами.
Ретротранспозоны залечивают двухнитевые разрывы ДНК. Обычно двухнитевой разрыв залечивается с помощью гомологичной молекулы ДНК, например, сестринской, только что реплицированной нити. Однако, если клетка лишена обычной системы залечивания двухнитевого разрыва, то в качестве заплатки может быть использована подвижная ДНК. Оказалось, что в роли такой подвижной ДНК может выступать реплицирующаяся ДНК ретротранспозонов. В этом случае спасательную функцию осуществляет класс ретротранспозонов. Заплатка позволит хромосоме сохранить целостность и не утратить концевого фрагмента. Правда, брешь в двухнитевой спирали ДНК будет залеплена заплаткой из ретротранспозона, то есть исходная нуклеотидная последовательность не будет восстановлена. Однако если район разрыва не содержал существенного гена, то клетка, а возможно, и организм сохранят жизнеспособность. Возможность участия ретротранспозонов, содержащих длинные концевые повторы, в процессе заживления двухнитевых разрывов была обнаружена в клетках дрожжей недавно, поэтому молекулярные механизмы обнаруженного явления остаются пока невыясненными.
Ретротранспозоны выступают как компоненты генома, спасающие хромосому от укорачивания. При воспроизведении ДНК перед клеточным делением синтез ДНК начинается с образования затравки РНК, поскольку фермент ДНК-полимераза способен только добавлять дезоксирибонуклеотидные звенья к 3'-концу полинуклеотидной цепи, но неспособен начинать синтез цепи ДНК. Затравка затем удаляется, и бреши застраиваются. Однако на одном из концов реплицирующихся молекул останется брешь, которую не удается заделать с помощью ДНК-полимеразы, работающей в 5'-3' направлении. Возникает опасность, что одиноко выступающий однонитевый конец ДНК будет уничтожен каким-либо ферментом, в результате чего молекула укоротится с конца. Если не принять соответствующих мер, то при каждом акте репликации ДНК хромосома будет укорачиваться с концов. В конечном итоге могут быть утрачены важные гены и клетка погибнет. В качестве спасателей в ряде случаев выступают ретротранспозоны, относящиеся к семействам, без длинных концевых повторов. Ретротранспозоны перемещаются, образуя повторяющуюся структуру, в которой элементы соединены друг с другом по типу «голова к хвосту». Таким образом, если эти ретротранспозоны и существовали когда-то как элементы-паразиты, то впоследствии геном хозяина приспособил их для выполнения столь важной функции, как сохранение концевых участков хромосом. Эти ретротранспозоны стали уже не эгоистами, а бесценными помощниками, спасающими хромосому от потери генов.
Естественный отбор способствует возникновению динамического баланса между положительными и отрицательными воздействиями мобильных элементов на приспособленность генома. Те видовые геномы, которые не способны противостоять вторжению генетических паразитов, неизбежно вытесняются из биосферы. Однако в процессе коэволюции большинство видов выработало компромиссную стратегию для защиты генома от неограниченного размножения мобильных элементов. Увеличение размера генома, например, ограничено оптимальным соотношением между ядром и объемом клетки, способным поддерживать установившийся темп метаболических процессов.
Поэтому лишь те мобильные элементы, которые приобрели отрицательную обратную связь на эффект дозы, сохранились в ходе эволюции, в то время как более агрессивные ДНК-паразиты исчезли.
Структура МГЭ
У прокариот выделяют два вида МГЭ.
Инсерционные последовательности (IS)
Инсерционная последовательность (IS, IS-элемент, англ. Insertion sequence) — короткий фрагмент ДНК, простой мобильный генетический элемент, имеющие на концах короткие (10–40 п. н.) схожие последовательности, расположенные в обратном порядке (инвертированные повторы).
Инсерционные последовательности обладают двумя важными характеристиками — они мало похожи на другие мобильные элементы (около 700—2500 нуклеотидов) и кодируют лишь белки, вовлеченные в процесс транспозиции (в отличие от транспозонов, кодирующих еще и некоторые вспомогательные гены, например, гены резистентности к антибиотикам). При транспозиции участок ДНК хозяина в сайте-мишени (5–9 п. н.) удваивается и окаймляет IS прямыми повторами, т. е. одинаково ориентированными (рис. 1).
Эти белки обычно представлены транспозазой, которая катализирует ферментативную реакцию, позволяющую IS элементу перемещаться, а также регуляторный белок, который стимулирует или ингибирует активность транспозиции.
Кодирующий район в IS элементе обычно фланкирован обращенными повторами.
IS-элементы могут быть частью сложных транспозонов.
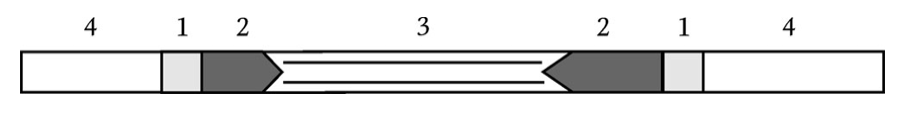
Рисунок 1 — Схема внедрения IS в ДНК хозяина: 1 – удвоенный сайт-мишень ДНК хозяина; 2 – инвертированные повторы; 3 – центральная область IS; 4 – ДНК клетки-хозяина
Бактериальная клетка обычно имеет множество IS. Наличие идентичных IS в различных бактериальных генетических структурах создает основу для их взаимодействия, интеграции и генетического обмена по принципу сайт-специфической рекомбинации.
Транспозоны (Tn) – более сложные структуры (2000–20 000 п. н.), в средней части имеющие различные гены, не обязательно связанные с транспозицией.
Tn с каждой стороны имеют плечи (или модули), которые являются IS, в прямой или инвертированной ориентации (рис. 2). Поскольку IS всегда имеют инвертированные повторы, то и Tn имеют их на своих краях. Вероятно, Tn возникли путем объединения двух первоначально независимых IS и области между ними. Многие транспозоны несут гены устойчивости к антибиотикам. Процесс транспозиции у бактерий включает дупликацию транспозона, при этом одна копия остается на старом месте, а другая возникает на новом.
Термин «транспозоны» в настоящее время обычно применяют ко всем МГЭ.

Рисунок 2 — Прямая (а) и инвертированная (б) ориентация плеч транспозонов
Структуры, аналогичные транспозонам бактерий, обнаружены у дрожжей, кукурузы, дрозофилы и других эукариотических организмов. Для них характерны типичные признаки транспозонов – инвертированные повторы и дупликация сайт-мишени. У дрожжей структуры Ty (Transposon yeast) встречаются в среднем в количестве 30–35 на клетку. Они имеют типичную структуру бактериального транспозона с плечами по 330 п. н. Такую же структуру имеют и самые первые из открытых МГЭ у кукурузы (около 5000 п. н.), получившие при открытии название «контролирующие элементы». У дрозофилы были обнаружены многочисленные транспозоноподобные структуры – МДГ (мобильные диспергированные гены). Одной из возможных функций МДГ является создание изолирующих барьеров для скрещивания на пути к видообразованию.
Все вирусы представляют собой «мобильные гены». Вирусы, плазмиды, транспозоны можно рассматривать в русле единого явления в эволюции генетической системы. Все эти структуры не только сами способны перемещаться по геному, но и могут «перетаскивать» соседние участки генома клетки-хозяина. Такое явление получило название трансдукции. Многообразие мобильных генетических структур послужило основой для гипотезы горизонтального переноса генов в эволюции.