

5.4. Уравнение продолжительности просаливания рыбы н.Н. Рулева
Продолжительность просаливания рыбы является важным технологическим параметром, необходимым для расчета технологического оборудования для посола рыбы.
При выводе этого уравнения Н.Н. Рулевым были сделаны следующие допущения:
пограничный слой у поверхности раздела фаз рыба-тузлук отсутствует, то есть движение соли в рыбу начинается непосредственно от границы раздела;
скорости внешней и внутренней диффузии равны;
на момент окончания посола имеет место состояние равновесия между фазами, то есть концентрация соли в тузлуке и в тканевом соке рыбы одинаковы.
В целом уравнение просаливания рыбы можно представить как массообмен между жидкостью (тузлуком) и твердым телом (рыбой). В твердых телах явление конвекции, как известно, отсутствует, поэтому внутренняя диффузия соли по своей сути аналогичная молекулярной диффузии. Уравнение массопроводности применительно к посолу рыбы можем представить в виде
![]() ,
(78)
,
(78)
где
![]() -
количество соли, проникшее в рыбу за
время
;
-
количество соли, проникшее в рыбу за
время
;
![]() - коэффициент
массопроводности рыбы, или коэффициент
диффузии соли в рыбе, м2/с;
- коэффициент
массопроводности рыбы, или коэффициент
диффузии соли в рыбе, м2/с;
- внешняя площадь поверхности рыбы, м2;
![]() - равновесная
концентрация соли в системе рыба-тузлук,
%;
- равновесная
концентрация соли в системе рыба-тузлук,
%;
- рабочая (текущая) концентрация соли в тканевом соке рыбы, %;
- средняя длина пути движения соли в рыбу, м.
Количество соли, проникшее в рыбу можно представить в следующем виде
![]() ,
(79)
,
(79)
где - концентрация соли, %;
- объем тканевого сока в рыбе, м3.
Если принять
допущение, что объем тканевого сока в
рыбе при посоле не изменяется, то есть
![]() ,
то
,
то
![]() .
(80)
.
(80)
Тогда после подстановки выражения (80) в уравнение (78) получим
![]() .
(81)
.
(81)
Объем тканевого сока , есть
![]() ,
(82)
,
(82)
где
![]() -
объем рыбы, м3;
-
объем рыбы, м3;
- содержание влаги в рыбе, доли единицы.
Тогда, после подстановки выражения (82) в уравнение (81) получим
![]() .
(83)
.
(83)
Разделим обе части
уравнения (83) на произведение
![]() ,
получим
,
получим
![]() ,
(84)
,
(84)
где
![]() -
удельная поверхность рыбы
,
м2/кг;
-
удельная поверхность рыбы
,
м2/кг;
![]() - скорость
просаливания;
- скорость
просаливания;
![]() - градиент
концентрации соли.
- градиент
концентрации соли.
Тогда, уравнение для скорости просаливания рыбы примет вид
![]() .
(85)
.
(85)
Если сделать допущение, что форма тела рыбы приближается к пластине, то суммарная площадь поверхности ее спинки и брюшка будет несоразмеримо мала по сравнению с площадью боковой поверхности тела. Следовательно, при определении полной площади поверхности тела рыбы суммарной площадью поверхности спинки и брюшка можно пренебречь.
При посоле в рыбе
имеет место двусторонняя симметричная
диффузия соли, движущая сила которой
![]() выражается через отношение разности
концентраций в наружном тузлуке
и внутри рыбы
к средней длине пути движения соли в
рыбе. Следовательно, значение
можно принять равным не ½ толщины рыбы,
имеющей форму пластины, а ¼ (рис. 11):
выражается через отношение разности
концентраций в наружном тузлуке
и внутри рыбы
к средней длине пути движения соли в
рыбе. Следовательно, значение
можно принять равным не ½ толщины рыбы,
имеющей форму пластины, а ¼ (рис. 11):
![]() ,
(86)
,
(86)
где - толщина рыбы, м.
δср.
δср.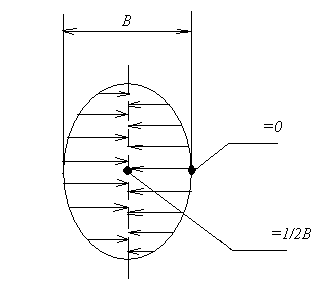


Рисунок 11 – Определение средней длины пути движения соли в рыбу
С учетом выражения (86), можно выразить толщину рыбы через ее удельную поверхность, как представлено ниже
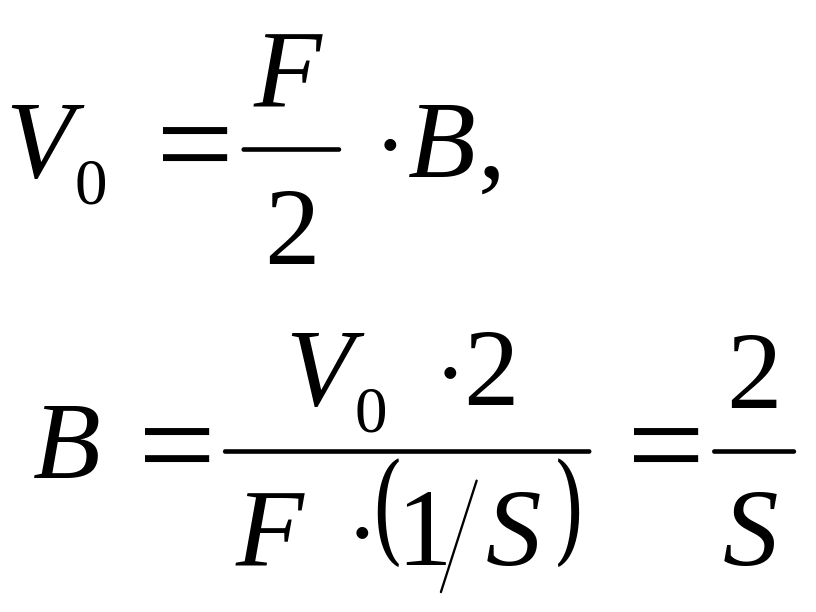 ,
(87)
,
(87)
где - объем рыбы, м3;
- полная площадь поверхности тела рыбы (без учета площади поверхности спинки и брюшка), м2;
- удельная поверхность рыбы, м2/кг.
Тогда для рыбы, имеющей тело в форме пластины, средняя длина пути соли в рыбу составит
![]() .
(88)
.
(88)
Подставив выражение (86) и (87) в уравнение для скорости просаливания рыбы (85), получим
![]() ,
(89)
,
(89)
![]() ,
(90)
,
(90)
![]() .
(91)
.
(91)
С помощью уравнений (90) и (91) определяют скорость просаливания рыбы, как с использованием ее толщины, так и с использованием удельной поверхности рыбы.
Полученные уравнения (90) и (91) позволяют проанализировать факторы, оказывающие наиболее значительное влияние на скорость просаливания, к этим факторам следует отнести:
- толщину рыбы или ее радиус, зависимость носит не только прямо пропорциональный, но и квадратичный характер;
- коэффициент диффузии, величина которого определяется химическим составом рыбы (содержанием в ней жира, чем оно больше, тем меньше величина при прочих равных условиях), а также температурой процесса (чем выше температура посола, тем больше величина коэффициента диффузии), характер зависимости – прямо пропорциональный;
- движущую силу процесса просаливания, определяемую разностью концентраций поваренной соли в тузлуке и в тканевом соке рыбы, причем следует отметить, что определяющее значение в этой разности имеет концентрация тузлука, которая определяется крепостью посола.
В уравнениях (90) и (91) коэффициент диффузии не зависит от геометрических характеристик продукта - толщины и удельной поверхности. Интегрируя указанные уравнения можно получить уравнения для определения продолжительности просаливания Н.Н. Рулева
![]() ,
(92)
,
(92)
![]() .
(93)
.
(93)
Постоянные интегрирования для уравнений (92) и (93) составляют
![]() ,
(94)
,
(94)
![]() .
(95)
.
(95)
Пользуясь уравнениями (92) и (93), Н.Н. Рулев определил значение коэффициентов диффузии для атлантических сельдевых рыб различной жирности при температуре посола от 5 до 10 °С. Рулевым была получена эмпирическая формула для определения коэффициента диффузии
![]() ,
см/сут.
,
см/сут.
Для учета температуры посола им был введен температурный коэффициент
![]() ,
(96)
,
(96)
где
![]() -
температурный коэффициент, или величина,
на которую изменяется коэффициент
диффузии в зависимости от температуры
посола (около 2-3 % на каждый 1 °С.
-
температурный коэффициент, или величина,
на которую изменяется коэффициент
диффузии в зависимости от температуры
посола (около 2-3 % на каждый 1 °С.
В целом погрешность формул (92) и (93) составляет 20 % что вполне допустимо в инженерных расчетах для физической величины, зависящей от таких неопределенных факторов, как химический состав и структура тканей рыбы.
Коэффициент диффузии в уравнениях (92) и (93) не всегда тождествен коэффициенту просаливания, что следует из допущений, принятых при выводе уравнения просаливания. Прежде всего, в отношении того, что отсутствует пограничный опресненный слой у поверхности раздела фаз рыба-тузлук и движение соли в рыбу начинается непосредственно от ее поверхности.
Согласно фронтальной теории Крина, такой пограничный слой существует, и в этом слое концентрация тузлука меньше равновесной в слое тузлука , причем значение изменяется в процессе диффузионных характеристик мышечной ткани рыбы за счет изменения нативных свойств белков под действием хлорида натрия.
Кроме того, коэффициенты просаливания, также как и коэффициенты диффузии изменяются по величине в процессе посола, а в формулах Рулева, их значение принято постоянным.
Коэффициент диффузии в формулах Рулева не учитывает погрешности математического моделирования, которые можно учесть коэффциентом просаливания (массопроводности).
Профессором Ершовым, профессором Глазуновым и инженером Димовой предложено выражение для определения солености в центре рыбы и в ее объеме в любой момент времени , исключающее погрешности формул Рулева
![]() ,
(97)
,
(97)
где
![]() -
толщина рыбы, м;
-
толщина рыбы, м;
- коэффициент диффузии соли, м2/с, для которого получена эмпирическая формула
![]() м2/с.
м2/с.
Лекция № 6: Изменения в соленой рыбе при хранении. Режимы и сроки хранения соленых гидробионтов. Вредители соленых продуктов водного сырья. Основные дефекты соленой рыбы
Наиболее существенным изменениям при посоле подвергается масса соленой рыбы. Она может, как увеличиваться в процесса просаливания, так и уменьшаться.
Выход части воды в системе рыба:тузлук — диффузионно-осмотический процесс. Леванидов И. П. отдавал предпочтение осмотической теории перемещения влаги, считая, что некоторые структурные элементы мышечной ткани рыб обладают свойствами полунепроницаемых мембран, поэтому разность осмотических давлений в клетке и межклеточном пространстве является основной движущей силой влагопереноса.
Осмотическое давление разбавленных растворов описывается уравнением Вант-Гоффа:
![]() ,
(98)
,
(98)
где
![]() —
изотонический коэффициент, равный для
хлорида натрия 2;
—
изотонический коэффициент, равный для
хлорида натрия 2;
![]() — молярная
концентрация растворенного вещества,
моль/л;
— молярная
концентрация растворенного вещества,
моль/л;
— универсальная газовая постоянная, Дж/(К·моль);
![]() — температура
раствора, К.
— температура
раствора, К.
Л. П.Миндер склонялся к тому, что влага из рыбы в основном извлекается засчет сил обычной молекулярной диффузии. Объясняя эту гипотезу тем, что мембраны у снулой рыбы из сплошных превращаются в сетчатые, которые проницаемы не только для воды, но и для органических веществ. Поэтому осмотические явления отсутствуют. Вероятно, оба процесса: осмос и диффузия имеют место при перемещении влаги из рыбы при ее просаливании.
Потеря влаги и поглощение соли мышечной тканью рыбы связаны обратнопропорциональной зависимостью. Таким образом, чем больше соли диффундировало в рыбу, тем меньше влаги в ней осталось. Следует отметить, что это справедливо при соленостях больше пяти процентов.
С другой стороны, в процессе хранения соленой рыбы в системе «рыба — тузлук» также имеют место массообменные процессы.
На I этапе хранения (до 25-30 суток в зависимости от солености рыбы, направляемой на посол) влага из рыбы удаляется. При последующем хранении (II этап) происходит впитывание влаги, так называемое набухание рыбы.
Процессы насыщения тканей рыбы водой в последующем хранении различные исследователи объясняют по-разному. Некоторые исследователи считают, что в результате гидролиза белков при посоле и последующем хранении происходит увеличение степени их дисперсности, что приводит к высвобождению связанной воды и ее диффузии из рыбы в тузлук (I этап). Размеры капилляров увеличиваются и уже не под действием осмотического давления, а за счет атмосферного давления происходит всасывание тузлука (II этап). Миндер Л. П. предполагает, что набухание вызвано двумя причинами: на первом этапе из рыбы успевает выйти больше воды, чем требуется для установления равновесия, следовательно, на втором этапе происходит поглощение тузлука (1 причина); пептизацией белков глобулинового характера в растворах низкой концентрации (2 причина). Отмечается также, что эти два процесса протекают одновременно и связаны между собой.
Представляет интерес следующее объяснение теории набухания. Перемещение воды в рыбу после завершения процесса влагоотдачи свидетельствует о нарушении электростатического равновесия в системе «рыба:тузлук». Это происходит за счет увеличения полярных групп при гидролизе белковых веществ и липидов и накопления заряженных аминных и карбоксильных групп и свободных аминокислот. Прекращение набухания при расщеплении белковых молекул в период созревания объясняется завершением процесса перераспределения слоев диполей воды около ионов Na+ и Cl–, причем, чем меньше расстояние между молекулами воды и ионами Na+ и Сl–, тем сильнее притяжение между ними. Набуханию более подвержено мясо нежирной рыбы, чем рыбы со значительным количеством жира в мышечной ткани.
Рассмотрим кинетику процессов извлечения воды из рыбы (I этап), которая характеризуется двумя периодами. В первом периоде происходит выделение межклеточного сока, который диффундирует через промежутки между волокнами. По мере выхода сока скорость выделения его в первый период уменьшается. Такое явление обычно происходит, когда концентрация соли в тканевом соке рыбы не превышает 12 %. После повышения концентрации соли во влаге рыбы выше 12 % солерастворимые белки тканей рыбы высаливаются, ткани сжимаются и уменьшаются в объеме. Вследствие этого увеличивается поперечное сечение промежутков между волокнами и уменьшается сопротивление выходу воды, влага выделяется более интенсивно, а скорость извлечения влаги уменьшается менее резко. Во второй период преимущественно извлекается внутриклеточный сок. Общее количество переместившейся воды из рыбы зависит от равновесной концентрации соли в ней.
На втором этапе больше влаги в рыбу в процессе ее хранения диффундирует при меньших концентрациях соли после посола. Это, видимо, можно объяснить тем, что при меньших концентрациях соли белки в меньшей степени высаливаются. Влага из клеток удаляется, в основном, за счет разности осмотического давления, при этом из рыбы удаляется больше воды, чем требуется для установления равновесия. На втором этапе хранения концентрация соли в клетке и тузлуке выравнивается за счет диффузии влаги в клетку опять-таки из-за разности осмотического давления. Эти процессы, по-видимому. протекают до полного выравнивания содержания соли как по всему объему рыбы, так и между рыбой и тузлуком. При увеличении концентрации соли во столько же раз увеличивается количество перемещающейся из рыбы воды. При постоянной концентрации соли в рыбе отношение количества перемещающейся воды к содержанию ее до просаливания является величиной постоянной, то есть чем больше начальное содержание воды в рыбе, тем больше ее выделяется при посоле. Скорость перемещения влаги из рыбы выше скорости перемещения соли в рыбу.
Если увеличивать температуру посола, то количество извлекаемой воды из рыбы при прочих равных условиях тоже увеличится.
Изменения массы рыбы при посоле обратимы не полностью. На эти изменения накладывают свой отпечаток начальные условия. Надо учитывать и изменения азотистых веществ и жира наряду с перемещениями воды и соли.
Потери плотных веществ при 0 °С больше, чем при 20 °С, но с увеличением плотности тузлука разница сглаживается. Это объясняется тем, что с повышением температуры усиливается высаливание белков, и потери плотных веществ меньше при теплом посоле, чем при холодном, а большую потерю массы при теплом посоле можно объяснить большей потерей воды.
Потери органических веществ в тузлуке при просаливании зависят и от продолжительности посола. При законченном посоле потери органических веществ выше, чем при прерванном посоле. Однако при дальнейшем хранении в тузлуке, потери органических веществ для рыбы законченного посола меньше, чем для рыбы прерванного посола.
Потери органических веществ у мороженой рыбы выше, чем у свежей. Такая закономерность особенно проявляется при низких температурах посола. По-видимому, это объясняется денатурационными изменениями белков, в результате чего нарушается связь белка и воды. Однако выход соленой продукции, приготовленной из мороженой рыбы, выше из3за меньших потерь воды.
Костные ткани в процессе посола также претерпевают изменения. Эти изменения зависят от минерализации костных тканей. Теоретически в случае абсолютно жесткой структуры кости при посоле должны увеличивать массу за счет насыщения солью воды, содержащейся в них. Однако при посоле тузлуком
(температура от 2 до 5 °С) у соленой трески масса костей увеличилась на 7,5%, а при посоле этой же трески в тузлуке с температурой от 7 до 13 °С масса костей уменьшилась на 1,5 %. Потери воды и плотных веществ, за исключением слабых посолов, больше, чем рыба приобретает соли. Поэтому масса соленой рыбы, как правило, меньше 100 %.
Тощие рыбы, как уже отмечалось, теряют в массе значительно больше, чем жирные. В свою очередь, чем крепче посол, тем больше рыба теряет воды и плотных веществ. Способ и техника посола также влияют на выход соленой рыбы.
Наибольший выход продукции дает смешанный посол, а наименьший — стоповый посол. При стоповом посоле нижние слои рыбы испытывают значительное давление, что ведет к уменьшению выхода рыбы. Влага, выделяющаяся из рыбы при стоповом посоле, свободно стекает, безвозвратно утрачиваясь. Также выход соленой рыбы уменьшается и при плотной ее укладке в таре.
Качество сырца также влияет на выход соленой рыбы. Рыба-сырец, посоленная в состоянии посмертного окоченения, теряет в массе меньше, чем рыба-сырец, находящаяся в стадии автолиза. При автолизе структура тканей рыхлая, соль при этом активно проникает к белкам и блокирует белковые молекулы, уменьшая их гидрофильность, вследствие этого большое количество влаги выделяется наружу.
Качество соли оказывает значительное влияние на выход соленой рыбы. Известно, что одновалентные металлы почти не вступают в связь с белками, поэтому Na+ не вступает во взаимодействие с белками рыбы. Двухвалентные ионы Са2+, Mg2+, которые могут встречаться в поваренной соли, наоборот, обладают высокой способностью вступать в соединение с белками, вызывая их денатурацию и уменьшая водоудерживающую способность. Если примесей в поваренной соли много, то выход продукции, обработанной такой солью, будет ниже.
Таким образом, на изменения массы и объема рыбы при посоле оказывают влияние следующие факторы: химический состав (жирность) рыбы, крепость тузлука и температура посола, способ и техника посола, механические воздействия, оказываемые на рыбу в процессе посола, качество сырья и качество используемой соли.
Процесс созревания — комплекс сложных превращений органических веществ рыбы, на который влияет значительный ряд изменяющихся параметров, таких как различный химический состав, строение и структура тканей, физические и другие воздействия, которые сопровождают приготовление и хранение соленой продукции. Зависимость созревания от множества изменяемых параметров не позволяет применить единую аналитическую оценку процесса.
В настоящее время известно два научных направления, в той или иной мере описывающее созревание соленых рыб.
Специалисты АтлантНИРО полагают об идентичности или незначительных отличиях степени гидролиза белка определенного строения пептидгидролазами пищеварительных органов разных рыб, то есть на скорость протеолиза прежде всего влияет количество и расположение гидролизуемых связей в белке при одном и том же наборе ферментов внутренностей. Другими словами, процесс созревания соленой рыбы — это комплекс ферментативных превращений белков, липидов и углеводов. Специфичность строения и состав субстратов влияют на качественный состав продуктов созревания, а активность ферментов и количество гидролизуемых связей в исходных продуктах — на скорость созревания (субстратная гипотеза).
Субстратная гипотеза получила дальнейшее развитие в лабораториях ТИНРО. Специалисты ТИНРО полагают, что роль и последовательность действия мышечных и пищеварительных ферментов влияют также на степень созревания соленой рыбы. Исследования показали, что продукты протеолиза соленых рыб, полученные под действием внутримышечных ферментов и под суммарным действием ферментов внутренностей, различаются не только количественно, но и имеют разную направленность. Например, в мясе целых рыб образуется большее количество низкомолекулярных продуктов: аминокислот и мелких пептидов, чем в мясе соленых разделанных рыб. В то же время изучение группового состава небелковых азотсодержащих соединений в течение первого периода хранения (месячного для сельди иваси и двухмесячного для тихоокеанской сельди) показало, что групповой состав идентичен для целых и разделанных рыб. Отсутствие существенных различий в количестве и составе продуктов протеолиза соленых разделанных и целых рыб позволяет предположить, что первоначально протеолиз протекает в основном под воздействием мышечных ферментов, а именно пептидгидролаз мышечной ткани. Отсюда следует, что процесс созревания зависит не от скорости проникновения ферментов внутренностей в мышечную ткань, а от степени подготовленности ее, которое лимитируется действием внутримышечных ферментов. На этой основе специалисты ТИНРО предложили разделить период созревания на три этапа: предсозревание (I этап), созревание (II этап), образование основных признаков соленой рыбы (III этап).
На первом этапе под воздействием пептидгидролаз мышечной ткани происходит незначительное накопление всех небелковых фракций, нарушается расположение полипептидных цепей внутри молекулы белка, что способствует образованию крупных пептидных фрагментов. Катепсин Д мышечной ткани воздействует на белки как типичная эдонтидаза и, вероятно, процесс подготовки белков к воздействию других ферментов, в том числе и ферментов внутренностей, зависит от катепсина Д.
Второй этап (созревание) характеризуется активно идущим протеолизом под суммарным воздействием ферментов мышечной ткани и внутренностей. Отмечается рост в количественном отношении азотсодержащих веществ и в большей мере — триптофансодержащих. Продукты распада белков первого этапа, а также те белки, которые оказались устойчивыми к действию внутримышечных ферментов, под действием ферментов пищеварительного тракта подвергаются разрушению до мелких пептидов и свободных аминокислот.
На третьем этапе происходит образование вкуса и аромата. Предположительно образование вкуса и аромата в соленой рыбе обусловлено реакциями синтеза между продуктами деградации компонентов мышечной ткани. Однако в настоящее время лишь косвенно подтверждаются эти предположения.
В образовании вкуса и аромата участвуют такие продукты расщепления липидов, как летучие карбонильные соединения и низкомолекулярные жирные кислоты. Большая часть из этих веществ обладает запахом и возможностью широкого взаимодействия с другими соединениями. Например, из сельди баночного посола выделено и идентифицировано 11 летучих кислот и 6 карбонильных соединений: изовалериановый, н-валериановый, лауриновый, уксусный, пропионовый и масляный, альдегиды.
Косвенное подтверждение взаимодействия продуктов расщепления белков и липидов найдено специальным определением так называемых оснований Шиффа. Основания Шиффа — это амины, которые образуются при реакции связывания малонового альдегида с двумя аминогруппами. Основания Шиффа обладают флюоресценцией с максимумом при длине волны 450 нм. Количество оснований Шиффа по мере хранения соленой рыбы возрастает, к тому же у хорошо созревающих рыб их больше.
На образование вкуса и аромата оказывают влияние и бактерии, например, молочнокислые, под действием которых образуются кислоты органического происхождения. Органические кислоты улучшают вкус и аромат соленой рыбы. Способность к созреванию мяса одного и того же вида рыбы, но в разные периоды годичного цикла, можно косвенно оценить по физиологическому состоянию рыбы и по кислотно-щелочному коэффициенту, который показывает соотношение между общим количеством соединений кислого и щелочного характера, образующихся при созревании соленой рыбы. Если у рыбы активность мышечных пептидгидролаз при pH от 6 до 6,5 более 4, то она имеет наибольшую способность к созреванию, при активности пептидгидролаз от 2 до 4 — менее созревающая рыба, а активность ферментов менее 2 — слабосозревающая рыба.
Определение активности мышечных пептидгидролаз свежих рыб при pH, оптимальном для действия катепсина Д (от 3,5 до 4) и катепсина С (при pH тканевого сока) позволяет оценить способность к созреванию той или иной рыбы.
Наиболее точно способность к созреванию можно оценить по комплексу показателей: глубине гидролиза белков (ГГБ); аминно-небелковому коэффициенту (К); разнице в оптической плотности (РОП) крупных пептидов (полипептидов), мелких пептидов (ди3 трипептидов), аминокислот и триптофана до и после термостатирования в буфере pH от 6 до 7.