

30. Инбридинг (случайный, неслучайный, тотальный) , его роль как фактор изменения генофонда популяции.
Спаривание родственных между собой самцов и самок называется инбридингом. Потомство, полученное в результате родственного подбора, называют инбредным. Спаривание неродственных животных называют аутбридингом. Родство между животными означает, что они имеют одного или нескольких общих предков. В результате этого инбредные животные имеют между собой и с родителями определенное генетическое сходство, в частности по составу аллелей в их генотипе. Поэтому в результате спаривания родственных, а следовательно, и сходных между собой животных у их потомства происходит накопление аллелей и генотипов того предка, который является общим для спариваемых родственных особей. Инбридинг приводит к повышению частоты гомозиготных генотипов у потомков и снижению частоты гетерозиготных генотипов при сохранении частот аллелей.
Родственное спаривание сопровождается снижением генетической изменчивости.
Наиболее близкородственной формой размножения в животном мире является самооплодотворение, встречающееся редко у низших филогенетических форм. У растений такой процесс прослеживается при самоопылении (например, у гороха, ячменя и др.). В практике животноводства применяют разные типы инбридинга, когда спаривают близких между собой родственников (отца с дочерью, мать с сыном, брата с сестрой); это называется кровосмешением. Другая группа подбора при инбридинге — это разведение в близком родстве, когда спариваются менее близкие родственники: полубрат — полусестра, бабушка — внук, внучка — дед, и еще более отдаленные родственники, то есть разведение в умеренном и отдаленном родстве.
В практике животноводства инбридинг применяют как при чистопородном разведении, так и при межпородном скрещивании. При этом степень родства между спариваемыми животными разнообразна.
В современной селекции приняты следующие схемы и терминология инбридинга:
-
родственное разведение - инбридинг;
-
использование родственного разведения в нескольких поколениях - ин-энд-инбридинг;
-
кровосмешение (близкородственное разведение) — клозебридинг;
-
спаривание животных из разных инбредных линий одной породы — инбредлайнкроссинг;
-
спаривание животных из близкородственных линий — стра-инкроссинг;
-
спаривание инбредных самцов с неинбредными самками — топкроссинг,
-
скрещивание инбредных самцов с неинбредными самками другой породы — топкроссбридинг;
-
скрещивание инбредных самцов одной породы с инбредными самками другой породы — инкроссбридинг;
-
спаривание инбредных маток с аутбредными самцами — боттомкроссинг.
Наиболее распространен в практике учет инбридинга, предложенный Шапоружем (1909). По этому методу учитывается число рядов поколений"; отделяющих потомка от предка, на которого осуществлен инбридинг. Начиная от ряда родительского поколения, обозначаемого римской цифрой I, далее каждый последующий ряд записывается как II, III, IV и т. д. Повторяющийся предок в родословной потомка со стороны матери и отца записывается с указанием ряда поколений, в которых он присутствует по материнской и отцовской стороне родословной.
Более точный метод определения степени инбридинга был разработан С. Райтом (1921) на основании принципа путевого анализа. Для этого была предложена формула коэффициента инбридинга. Позднее профессором Д. А. Кисловским в формулу Райта было внесено уточнение, что делает вычисление более удобным.
Формула Райта — Кисловского представляет коэффициент инбридинга F в виде дроби в границах от 0 до 1 (или в %).
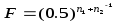
Чем больше величина F приближается к единице (или 100%), тем сильнее инбридирован потомок на предка, тем больше у потомков можно ожидать проявления инбредной депрессии и тем более вероятность повышения гомозиготности потомка по генам предка. Коэффициент инбридинга не указывает в абсолютных цифрах или в процентах, насколько гомозиготен потомок; он только свидетельствует о вероятности того, насколько примененный родственный подбор увеличит гомози-готность потомка по сравнению с исходным состоянием генотипа.
У животных с высокой степенью гетерозиготности инбридинг усиливает гомозиготность значительно быстрее, чем у Животных, уже имевших значительную гомозиготность до инбридинга.
Уровень гомозиготности потомка зависит от того, были ли его родители гетерозиготны или гомозиготны.
Использование гетерозиготных производителей — наиболее перспективный вариант инбридинга при проведении племенной работы в стаде, так как он не сопровождается депрессией у потомка, но обеспечивает более стабильное сходство с ценным предком.
Формула, предложенная Райтом для вычисления степени возрастания генетического сходства между двумя сравниваемыми животными, такова:
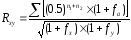
где: Rxy – коэффициент генетического сходства между животными x и y; n1 и n2 число поколений от данных животных до общего предка по женской и мужской стороне родословной; fx и fyкоэффициент возрастания гомозиготности для животных x и y; fa – тот же коэффициент для общего инбридированного предка.
Так, в XVIII в. в Англии братья Коллинги в результате селекции получили быка Комета шортгорнской породы, который был инбридирован на четырех выдающихся предков: быков Фаворита и Фольджамба и коров Феникс и Леди Майнард, при этом корова Леди Майнард и бык Фольджамб были инбредными животными. Коэффициент инбридинга был достаточно высоким (F=46,87%), но, несмотря на это, Комет сыграл важную роль в совершенствовании породы.
Депрессия может наблюдаться и при так называемом ложном инбридинге, который является следствием содержания животных в ряде поколений в одинаковых, часто изнеживающих условиях. В целях устранения депрессии при истинном и ложном инбридинге целесообразно периодически содержать животных, намеченных к спариванию, в различных условиях, что способствует формированию некоторого биологического несходства гамет самцов и самок. Этот прием называют интербридингом.
Использование инбридинга в селекции растений. Раздельнополость и приспособленность к перекрестному опылению, столь распространенные в растительном мире, и существование у растений многочисленных препятствий к самоопылению выработаны в процессе эволюции. Они не могли бы так широко распространиться, если перекрестное опыление не имело бы в борьбе за жизнь каких-то вполне определенных преимуществ перед самоопылением и не было бы, поэтому подхвачено естественным отбором. Естественным отбором создана и приспособленность растений определенных видов к соответствующим насекомым — их опылителям и, наоборот, насекомых определенных видов к соответствующим растениям. У ветроопыляемых же растений эволюционно развилась необходимость вырабатывать огромный избыток пыльцы; только тогда при случайных порывах ветра на рыльца пестиков попадет какая-то ее часть. К такому выводу Ч. Дарвин пришел на основании изучения природы. Для подтверждения его Дарвин поставил специальные опыты, продолжавшиеся более 10 лет на растениях 57 различных видов. Растения каждого вида он разделил на две разные части и представителей одной из них размножал в ряде последующих поколений путем самоопыления, а другой — путем перекрестного опыления. Дочерние растения выращивали в возможно более сходных условиях — в одних горшках или на одних грядках. У полученных от перекрестного опыления растений показатели высоты, веса и плодовитости оказались в среднем значительно больше. Так, высота растений, происшедших от самоопыления, по сравнению с полученными от перекрестного опыления составляла в среднем только 87%, а вес — лишь 67%.