

Ответы к зачету по цитологии
Билет № 37. Липиды и их назначение.
Ф-ции липидов:
Одна из основных функций жиров (липидов) – энергетическая. В ходе расщепления 1 г жиров до диоксида углерода и воды освобождается большое количество энергии – 38,9 кДж. Содержание жира в клетке составляет 5-15% массы сухого вещества. В клетках жировой ткани количество жира составляет до 90%. Накапливаясь в клетках жировой ткани животных, в семенах и плодах растений, жир служит запасным источником энергии.
Липиды выполняют т строительную функцию: они входят в состав клеточных мембран
Вследствие плохой теплопроводности жир способен выполнять функцию теплоизолятора. У некоторых животных (тюлени, киты) он откладывается в подкожной жировой ткани, которая у китов образует слой толщиной до 1 м.
Образование некоторых липоидов предшествует синтезу ряда гормонов. Следовательно, этим веществам присуща и функция регуляции обменных процессов.
Защитная (амортизационная).Толстый слой жира защищает внутренние органы многих животных от повреждений при ударах (например, сивучи при массе до тонны могут прыгать на каменистый берег со скал высотой 4-5 м).
Увеличения плавучести. Самые разные организмы — от диатомовых водорослей до акул — используют резервные запасы жира как средство снижения среднего удельного веса тела и, таким образом, увеличения плавучести. Это позволяет снизить расходы энергии на удержание в толще воды.
Билет № 38. История открытия нуклеиновых кислот.
В 1847 из экстракта мышц быка было выделено вещество, которое получило название «инозиновая кислота». Это соединение стало первым изученным нуклеотидом. В течение последующих десятилетий были установлены детали его химического строения. В частности, было показано, что инозиновая кислота является рибозид-5'-фосфатом, и содержит N-гликозидную связь.
В 1868 году швейцарским химиком Фридрихом Мишером при изучении некоторых биологических субстанций было открыто неизвестное ранее вещество. Вещество содержало фосфор и не разлагалось под действием протеолитических ферментов. Также оно обладало выраженными кислотными свойствами. Вещество было названо «нуклеином». Соединению была приписана брутто-формула C29H49N9O22P3.
Уилсон обратил внимание на практическую идентичность химического состава «нуклеина» и открытого незадолго до этого «хроматина» — главного компонента хромосом. Было выдвинуто предположение об особой роли «нуклеина» в передаче наследственной информации.
В 1889 г Рихард Альтман ввел термин «нуклеиновая кислота», а также разработал удобный способ получения нуклеиновых кислот, не содержащих белковых примесей.
Левин и Жакоб, изучая продукты щелочного гидролиза нуклеиновых кислот, выделили их основные составляющие — нуклеотиды и нуклеозиды, а также предложили адекватные структурные формулы, описывающие их свойства.
В 1921 году Левин выдвинул гипотезу «тетрануклеотидной структуры ДНК», оказавшуюся впоследствии ошибочной.
В 1935 году Клейн и Танхаузер с помощью фермента фосфатазы провели мягкое фрагментирование ДНК, в результате чего были получены в кристаллическом состоянии четыре ДНК-образующих нуклеотида. Это открыло новые возможности для установления структуры этих соединений.
В 1940-е годы научная группа в Кембридже под руководством Александера Тодда проводит широкие синтетические исследования в области химии нуклеотидов и нуклеозидов. В результате их работы были установлены все детали химического строения и стереохимии нуклеотидов. За цикл работ в этой области Александер Тодд был награждён Нобелевской премией в области химии в 1957 году.
Чаргаффом было установлена закономерность содержания в нуклеиновых кислотах нуклеотидов разных типов, получившая впоследствии название Правило Чаргаффа.
Открытие нуклеиновых кислот связано с именем молодого врача из города Базеля (Швейцария) Фридриха Мишера. После окончания медицинского факультета Мишер был послан для усовершенствования и работы над диссертацией в Тюбинген (Германия) в физиолого-химическую лабораторию, возглавляемую Ф. Гоппе-Зейлером. Тюбингенская лаборатория в то время была известна ученому миру. Пройдя практику по органической химии, Мишер приступил к работе в биохимической лаборатории. Ему было поручено заняться изучением химического состава гноя. Молодой ученый не возражал против предложенной темы, так как считал лейкоциты, присутствующие в гное, одними из самых простых клеток.
Путём многочисленных опытов он получил из гнойных клеток вещество ядерного происхождения. Мишер был уверен именно в ядерном его источнике. Поэтому он начал более тщательное выделение ядер. В то время еще никто в биохимических лабораториях не пытался выделить ядра или какие-либо другие субклеточные компоненты, так что и здесь он был пионером.
Продолжив дальше очищать ядро от других клеточных фрагментов, он получил странное вещество. Оно не разлагалось протеолитическими ферментами, значит, не являлось белком. Отсутствие растворимости в горячем спирте указывало на то, что это вещество не являлось и фосфолипидом. По-видимому, оно относилось к новому классу биохимических соединений.
Но Мишер с большой горячностью настаивал на точности своих результатов и добивался разрешения опубликовать их в печати. Тогда Гоппе-Зейлер решил проверить данные Мишера лично. Он и два его ассистента (одним из них был русский химик Любавин) в течение года шаг за шагом прошли все этапы аналитической работы Мишера и полностью подтвердили его данные, выделив нуклеин из клеток крови и из дрожжей.
В 1871 г. работа Мишера вместе с подтверждающими ее контрольными работами Гоппе-Зейлера и его ассистентов увидела свет. Существование нуклеина как специфического ядерного вещества стало научным фактом. Вскоре методика Мишера была применена для выделения нуклеина из различных тканей.
Термин «нуклеиновые кислоты» был предложен в 1889: нуклеиновыми они были названы потому, что впервые были открыты в ядрах клеток, а кислотами -- из-за наличия в их составе остатков фосфорной кислоты. Позже было показано, что нуклеиновые кислоты построены из большого числа нуклеотидов (от нескольких десятков до сотен миллионов). В состав каждого нуклеотида входит азотистое основание, углевод (пентоза) и фосфорная кислота.
Билет № 40.Биополимер ДНК и его мономеры. 4 типа дезоксирибонуклеотидов.
ДНК- это биологический полимер, состоящий из двух полинуклеотидных цепей, соединенных друг с другом. Мономеры – нуклеотиды, составляющие каждую из цепей ДНК, представляют собой сложные органические соединения. ДНК состоит из четырех азотистых оснований: производных пуринов – аденина (А) и гуанина (Г) и производных пиримидинов – цитозина (Ц) и тимина (Т), пятиатомного сахара пентозы – дезоксирибозы, а также остатка фосфорной кислоты.
Билет № 41. Первичная и вторичная структуры ДНК, комплементарность азотистых оснований.
Первичная структура нуклеиновых кислот – это нуклеотидный состав и определенная последовательность нуклеотидных звеньев в полимерной цепи.
Вторичная структура ДНК: В 1953 г. Дж. Уотсоном и Ф. Криком была предложена модель пространственной структуры ДНК. Согласно этой модели, молекула ДНК имеет форму спирали, образованную двумя полинуклеотидными цепями, закрученными относительно друг друга и вокруг общей оси. Двойная спираль правозакрученная, полинуклеотиды цепи в ней антипараллельны, т.е. если одна из них ориентирована в направлении 3'→5', то вторая - в направлении 5'→3'. Поэтому на каждом из концов молекулы ДНК расположены 5'-конец одной цепи и 3'-конец другой цепи. представляет собой две параллельные неразветвленные полинуклеотидные цепи, закрученные вокруг общей оси в двойную спираль.
Т акая
пространственная структура удерживается
множеством водородных связей, образуемых
азотистыми основаниями, направленными
внутрь спирали.
акая
пространственная структура удерживается
множеством водородных связей, образуемых
азотистыми основаниями, направленными
внутрь спирали.
Водородные связи возникают между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи. Эти основания составляют комплементарные пары (от лат. complementum - дополнение).
В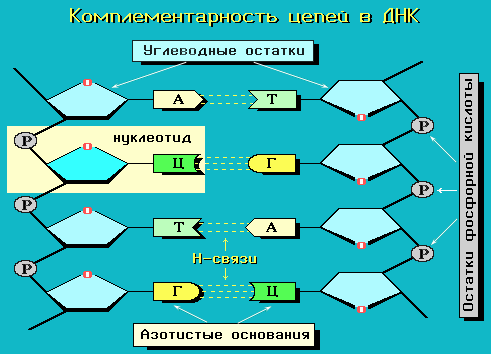 каждой цепи нуклеотиды соединяются
между собой ковалентными связями:
дезоксирибоза одного нуклеотида
соединяется с остатком фосфорной кислоты
последующего нуклеотида. Две цепи
объединяются в единую молекулу водородными
связями, возникающими между азотистыми
основаниями, входящими в состав
нуклеотидов, образующих разные цепи.
Пространственная конфигурация азотистых
оснований различна и количество таких
связей между разными азотистыми
основаниями неодинаково. Вследствие
этого они могут соединяться только
попарно: азотистое основание аденин
(А) одной цепочки полинуклеотида всегда
связано двумя водородными связями с
тимином (Т) другой цепи, а гуанин (Г) –
тремя водородными связями с азотистым
основанием цитозином (Ц) противоположной
полинуклеотидной цепочки. Такая
способность к избирательному соединению
нуклеотидов, в результате которого
формируются пары А-Т и Г-Ц, называется
комплементарностью.
Если известна последовательность
оснований в одной цепи (например,
Т-Ц-А-Т-Г), то благодаря принципу
комплементарности (дополнительности)
станет известна и противоположная
последовательность оснований (А-Г-Т-А-Ц).
каждой цепи нуклеотиды соединяются
между собой ковалентными связями:
дезоксирибоза одного нуклеотида
соединяется с остатком фосфорной кислоты
последующего нуклеотида. Две цепи
объединяются в единую молекулу водородными
связями, возникающими между азотистыми
основаниями, входящими в состав
нуклеотидов, образующих разные цепи.
Пространственная конфигурация азотистых
оснований различна и количество таких
связей между разными азотистыми
основаниями неодинаково. Вследствие
этого они могут соединяться только
попарно: азотистое основание аденин
(А) одной цепочки полинуклеотида всегда
связано двумя водородными связями с
тимином (Т) другой цепи, а гуанин (Г) –
тремя водородными связями с азотистым
основанием цитозином (Ц) противоположной
полинуклеотидной цепочки. Такая
способность к избирательному соединению
нуклеотидов, в результате которого
формируются пары А-Т и Г-Ц, называется
комплементарностью.
Если известна последовательность
оснований в одной цепи (например,
Т-Ц-А-Т-Г), то благодаря принципу
комплементарности (дополнительности)
станет известна и противоположная
последовательность оснований (А-Г-Т-А-Ц).
Цепи нуклеотидов образуют правозакрученные объемные спирали по 10 оснований в каждом витке. Последовательность соединения нуклеотидов одной цепи противополодна такой в другой цепи, т.е. цепи составляющие одну молекулу ДНК, разнонаправлены, или антипараллельны: последовательность межнуклеотидных связей в двух цепях направлена в противоположные стороны: 5'-3' и 3'-5'. Сахаро-фосфатные группировки нуклеотидов находятся снаружи, а азотистые основания внутри. Цепи, закрученные друг относительно друга, а также вокруг общей оси, образуют двойную спираль. Такая структура молекулы поддерживается в основном водородными связями.
Билет № 42. Третичная структура и структуры более высокого порядка.
Третичная структура ДНК (суперспирализация ДНК)
Каждая молекула ДНК упакована в отдельную хромосому. В диплоидных клетках человека содержится 46 хромосом. Общая длина ДНК всех хромосом клетки составляет 1,74 м, но она упакована в ядре, диаметр которого в миллионы раз меньше. Чтобы расположить ДНК в ядре клетки, должна быть сформирована очень компактная структура. Компактизация и суперспирализация ДНК осуществляются с помощью разнообразных белков, взаимодействующих с определёнными последовательностями в структуре ДНК. Все связывающиеся с ДНК эукариотов белки можно разделить на 2 группы: гистоновые и негистоновые белки. Комплекс белков с ядерной ДНК клеток называют хроматином.
Гистоны - белки с молекулярной массой 11-21 кД, содержащие много остатков аргинина и лизина. Благодаря положительному заряду гистоны образуют ионные связи с отрицательно заряженными фосфатными группами, расположенными на внешней стороне двойной спирали ДНК.
Существует 5 типов гистонов. Четыре гистона Н2А, Н2В, НЗ и Н4 образуют октамерный белковый комплекс (Н2А, Н2В, НЗ, Н4)2, который называют "нуклеосомный кор". Молекула ДНК "накручивается" на поверхность гистонового октамера, совершая 1,75 оборота (около 146 пар нуклеотидов). Такой комплекс гистоновых белков с ДНК служит основной структурной единицей хроматина, её называют "нуклеосома". ДНК, связывающую нуклеосомные частицы, называют линкерной ДНК. В среднем линкерная ДНК составляет 60 пар нуклеотидных остатков. Молекулы гистона H1 связываются с ДНК в межнуклеосомных участках (линкерных последовательностях) и защищают эти участки от действия нуклеаз.
В ядре каждой клетки присутствует около 60 млн молекул каждого типа гистонов.
Негистоновые белки хроматина
В ядре эукариотической клетки присутствуют сотни самых разнообразных ДНК-связывающих негистоновых белков. К негистоновым белкам принадлежат ферменты репликации, транскрипции и репарации. При участии структурных, регуляторных белков и ферментов, участвующих в синтезе ДНК и РНК, нить нуклеосом преобразуется в высококонденсированный комплекс белков и ДНК. Образованная структура в 10 000 раз короче исходной молекулы ДНК.
Билет № 43. Модель строения молекулы ДНК. Антипараллельность. Параметры двойной спирали, полный оборот (шаг) спирали.
Cоглатно этой модели молекула ДНК состоит из 2 антипараллельных полинуклеотидных цепей, образующих правозакрученную спираль, удерживаемую взаимодействием пар азотистых оснований в соответствии с правилами комплементарности, модель предложена Дж.Уотсоном и Ф. Криком в 1953, они же определили количественные характеристики двойной спирали ДНК. Диаметр спирали составляет 2нм, длина шага – 3-4нм. В каждый виток входит 10 пар нуклеотидов.
Билет№ 44. Локализация ДНК: хромосомная и экстрахромосомная, функции ДНК
Функция ДНК — хранение и передача наследственной информации.
Репликация (редупликация) ДНК
Репликация ДНК — процесс самоудвоения, главное свойство молекулы ДНК. Репликация относится к категории реакций матричного синтеза, идет с участием ферментов. Под действием ферментов молекула ДНК раскручивается, и около каждой цепи, выступающей в роли матрицы, по принципам комплементарности и антипараллельности достраивается новая цепь. Таким образом, в каждой дочерней ДНК одна цепь является материнской, а вторая — вновь синтезированной. Такой способ синтеза называется полуконсервативным.
Репликация происходит перед делением клетки. Благодаря этой способности ДНК осуществляется передача наследственной информации от материнской клетки дочерним.
Репарация («ремонт»)
Репарацией называется процесс устранения повреждений нуклеотидной последовательности ДНК. Осуществляется особыми ферментными системами клетки (ферменты репарации). В процессе восстановления структуры ДНК можно выделить следующие этапы: 1) ДНК-репарирующие нуклеазы распознают и удаляют поврежденный участок, в результате чего в цепи ДНК образуется брешь; 2) ДНК-полимераза заполняет эту брешь, копируя информацию со второй («хорошей») цепи; 3) ДНК-лигаза «сшивает» нуклеотиды, завершая репарацию.
Наиболее изучены три механизма репарации: 1) фоторепарация, 2) эксцизная, или дорепликативная, репарация, 3) пострепликативная репарация.
Изменения структуры ДНК происходят в клетке постоянно под действием реакционно-способных метаболитов, ультрафиолетового излучения, тяжелых металлов и их солей и др. Поэтому дефекты систем репарации повышают скорость мутационных процессов, являются причиной наследственных заболеваний (пигментная ксеродерма, прогерия и др.)
Билет№ 45. Строение и состав рнк.
РНК — полимер, мономерами которой являются рибонуклеотиды. В отличие от ДНК, РНК образована не двумя, а одной полинуклеотидной цепочкой (исключение — некоторые РНК-содержащие вирусы имеют двухцепочечную РНК). Нуклеотиды РНК способны образовывать водородные связи между собой. Цепи РНК значительно короче цепей ДНК.
Мономер РНК — нуклеотид (рибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания РНК также относятся к классам пиримидинов и пуринов.
Пиримидиновые основания РНК — урацил, цитозин, пуриновые основания — аденин и гуанин. Моносахарид нуклеотида РНК представлен рибозой.
Биологическая роль РНК связана преимущественно с синтезом белка, т. е. с реализацией наследственной информации. Именно РНК является посредником между ДНК и строящейся в клетке белковой молекулой.
Билет№ 46. Типы рнк. Информационная рнк, химический состав, структура, роль в клетке.
Выделяют три вида РНК: 1) информационная (матричная) РНК — иРНК (мРНК), 2) транспортная РНК — тРНК, 3) рибосомная РНК — рРНК.
Все виды РНК представляют собой неразветвленные полинуклеотиды, имеют специфическую пространственную конформацию и принимают участие в процессах синтеза белка. Информация о строении всех видов РНК хранится в ДНК. Процесс синтеза РНК на матрице ДНК называется транскрипцией.
Информационные РНК разнообразны по содержанию нуклеотидов и молекулярной массе (от 50 000 до 4 000 000). На долю иРНК приходится до 5% от общего содержания РНК в клетке. Функции иРНК: 1) перенос генетической информации от ДНК к рибосомам, 2) матрица для синтеза молекулы белка, 3) определение аминокислотной последовательности первичной структуры белковой молекулы.
Билет № 47. Рибосомальная рнк, состав, структура, формирование субъединицы хромосом.
Рибосомные рибонуклеиновые кислоты (рРНК) — несколько молекул РНК, составляющих основу рибосомы. Основной функцией рРНК является осуществление процесса трансляции — считывания информации с мРНК при помощи адапторных молекул тРНК и катализ образования пептидных связей между присоединёнными к тРНК аминокислотами.
Рибосомные РНК содержат 3000–5000 нуклеотидов; молекулярная масса — 1 000 000–1 500 000. На долю рРНК приходится 80–85% от общего содержания РНК в клетке. В комплексе с рибосомными белками рРНК образует рибосомы — органоиды, осуществляющие синтез белка. В эукариотических клетках синтез рРНК происходит в ядрышках. Функции рРНК: 1) необходимый структурный компонент рибосом и, таким образом, обеспечение функционирования рибосом; 2) обеспечение взаимодействия рибосомы и тРНК; 3) первоначальное связывание рибосомы и кодона-инициатора иРНК и определение рамки считывания, 4) формирование активного центра рибосомы.
Транскрипция эукариотических рибосомных генов осуществляется РНК-полимеразой I и происходит в ядрышках. Синтезируемый предшественник рРНК не подвергается ни кэпированию ни полиаденилированию. Установлено, что формирование ядрышек непосредственно связано с транскрипцией рибосомных генов ( Oaks и сотр., 1993 ). Сформировавшись, ядрышки становятся местом транспорта рибосомных белков и малых ядерных рибонуклеопротеинов ( мяРНП ). Хорошо видимая "гранулярная" зона в ядрышках содержит частично собранные рибосомы.