

2. Передача сигналов через внутриклеточные рецепторы
Стероидные и тиреоидные гормоны связываются с рецепторами внутри клетки и регулируют скорость транскрипции специфических генов. В отсутствие гормона внутриклеточные рецепторы связаны обычно с другими белками в цитозоле или ядре. Например, рецепторы глюкокортикоидов образуют в цитозоле комплекс с шапероном, что препятствует связыванию рецептора с молекулой ДНК. Взаимодействие гормона с центром связывания на С-концевом участке полипептидной цепи рецептора вызывает конформационные изменения и освобождение рецептора от шаперона. Происходит объединение 2 молекул рецептора с образованием гомодимера. Димер рецептора узнаёт специфическую последовательность нуклеотидов, которая расположена в промоторной области гена. Взаимодействие со специфическим участком ДНК HRE (от англ, hormone response element, элемент, реагирующий на воздействие гормона) обеспечивает центральный домен рецептора. Этот домен содержит аминокислотную последовательность, образующую 2 "цинковых пальца". В каждом "цинковом пальце" атом цинка связан с 4 остатками цистеина.
В структуре одного "цинкового пальца" имеется последовательность аминокислот, отвечающая за связывание с ДНК, а второй "цинковый палец" содержит последовательность аминокислот, участвующую в димеризации рецепторов. Взаимодействие комплекса гормон-рецептор с определённой последовательностью нуклеотидрв в промоторной части ДНК приводит к активации транскрипции. Рецепторы тиреоидных гормонов всегда связаны с ДНК. В отсутствие гормонов соответствующие рецепторы ингибируют экспрессию генов. Напротив, взаимодействие с гормоном превращает их в активаторы транскрипции.
3. Передача сигналов через рецепторы, сопряжённые с ионными каналами
Р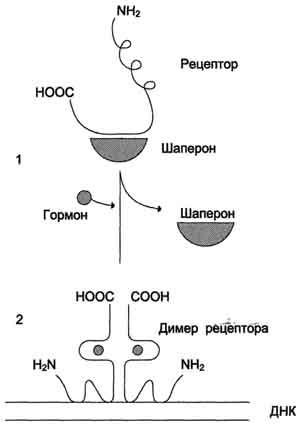 ецепторы,
сопряжённые с ионными каналами, являются
интегральными мембранными белками,
состоящими из нескольких субъединиц.
Они действуют одновременно как ионные
каналы и как рецепторы, которые способны
специфически связывать с внешней стороны
эффектор, изменяющий их ионную
проводимость. Эффекторами такого типа
могут быть гормоны и нейромедиаторы.
Известны рецепторы для ряда гормонов,
ассоциированных с ионными каналами, и
большинства медиаторов, среди которых
наиболее изучен рецептор ацетилхолина.
Рецептор ацетилхолина состоит из пяти
цилиндрообразных субъединиц, расположенных
в мембране параллельно друг другу:α2,
β, γ, δ. Между ними вдоль оси цилиндров
находится заполненный молекулами воды
канал. Каждая субъединица рецептора
состоит из большого количества гидрофобных
аминокислотных остатков.
ецепторы,
сопряжённые с ионными каналами, являются
интегральными мембранными белками,
состоящими из нескольких субъединиц.
Они действуют одновременно как ионные
каналы и как рецепторы, которые способны
специфически связывать с внешней стороны
эффектор, изменяющий их ионную
проводимость. Эффекторами такого типа
могут быть гормоны и нейромедиаторы.
Известны рецепторы для ряда гормонов,
ассоциированных с ионными каналами, и
большинства медиаторов, среди которых
наиболее изучен рецептор ацетилхолина.
Рецептор ацетилхолина состоит из пяти
цилиндрообразных субъединиц, расположенных
в мембране параллельно друг другу:α2,
β, γ, δ. Между ними вдоль оси цилиндров
находится заполненный молекулами воды
канал. Каждая субъединица рецептора
состоит из большого количества гидрофобных
аминокислотных остатков.
Рис. 11-6. Регуляция активности рецептора стероидных гормонов. 1 - в отсутствие гормона рецептор через гор-монсвязывающий домен образует комплекс с шапероном, что препятствует связыванию рецептора с молекулой ДНК; 2 - в присутствии гормона рецептор освобождается от шаперона, образуется димер рецептора, который присоединяется к молекуле ДНК и вызывает активацию транскрипции.
Кроме этого, все субъединицы содержат один спирализованный трансмембранный фрагмент, аминокислотные радикалы которого (полярные незаряженные аминокислотные остатки, в основном серии и треонин) выстилают центральный канал рецептора изнутри. В средней части субъединиц, обращённой к каналу, локализованы остатки лейцина. В присутствии ацетилхолина боковые взаимодействия между субъединицами поддерживают канал в открытом состоянии и создают возможность для транспорта ионов. В отсутствие ацетилхолина в результате изменения ориентации субъединиц относительно друг друга канал закрывается, так как выступающие внутрь канала остатки лейцина образуют плотное гидрофобное кольцо, блокируя движение гидратированных ионов в этой области.