45. Общая схема транспорта электронов в фотосинтетической этц растений. Структура и работа пластоцианинов.
Работа пластоцианинов (ПЦ)
Перенос электронов на ПЦ
Медь-содержащий (потому голубой) белок фотосинтетической ЭТЦ.
Имеет мономерную структуру (10,5 кДа), всего 99 аминокислот.
ПЦ-Cu2+ может восстанавливаться цитохромом f до ПЦ-Cu+
ПЦ-Cu+ диффундирует внутри лумена и связывается с P700+.
P700+ окисляет ПЦ-Cu+ обратно до ПЦ-Cu2+.
46. Общая схема транспорта электронов по фотосистеме-I. Строение железо-серных белков.
ФС-1 содержит 110 кофакторов (обычно «делимых» функционально-структурных частей).
ССК-1 состоит из молекул хлорофилла и каротиноидов, удерживаемых вместе при помощи 2 белков. Кол-во молекул пигментов варьирует у разных видов от 25 до 120.
Димеры хл. а образуют реакционный
центр Р700.
Перенос электрона от Р700 на первичный акцептор в ФС-1, которым является модифицированный хлорофилл А0 (хлорофилл А0 - этиомер по положению 13 хлоринового кольца)
7. . . . от хлорофилла А0 на филлохинон А1 (необязательный этап):
8.Перенос электрона от хлорофилла А0 или филлохинона А1 на железо-серные белки
Железо-серные белки (Fe-S-белки):
образуют комплекс из 3 белков Fx, Fb и Fa (А2)
последовательно передающий электрон от
Fx к Fа и затем на Fb. Железо (переходный металл) удерживается остатками цистеина.
9. Fb восстанавливает ферредоксин (ФД).
(ФД – белки, состоящие приблизительно из 1000 АК.
Структура - β(2)-α-β(2), которая включает путидаредоксин, терпредоксин и адренодоксин.
Содержит консервативный участок,образующий железо-серный кластер.
47. Общая схема транспорта электронов по фотосистеме-I. Строение и особенности функционирования ферредоксин-НАДФ+-редуктазы.
Fb восстанавливает ферредоксин (ФД).(ФД – белки, состоящие приблизит
ельно из 1000 АК. Структура - β(2)-α-β(2), которая включает путидаредоксин, терпредоксин и адренодоксин. Содержит консервативный участок,образующий железо-серный кластер.
. Ферредоксин-НАДФ+-редуктаза (ФНР)переносит электрон от восстановленного ФД на
НАДФ+. Этот же фермент может работать в обратном направлении и принимать электрон от НАДФН.
«Классическая» работа ФНР:
2 восстановленных ФД + НАДФ+ + H+ = 2 окисленных ФД + НАДФH 3 субстрата ФНР: (1) восст. ФД, (2) НАДФ+ и (3) H+.
Фермент имеет 2 кофактора: ФАД и флавин.
Растительная ФНР имеет два структурных домена: 1 домен – на N-конце «антипараллельный бета-баррель», связывающий ФАД; 2 – домен – на С-конце – «альфа-спираль – бета-цепь», связывающая НАДФ+. Активный центр фермента образуется обоими доменами.
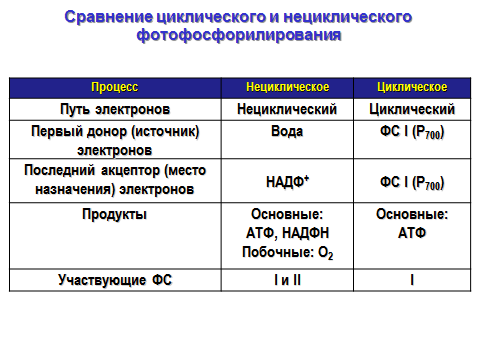
48. Сравнение циклического и нециклического фотофосфорилирования
Кооперация между двумя циклами связана не только с перекачкой СО2. Для восстановления ФГК в цикле Кальвина необходимы АТФ и НАДФН. Агранальные хлоропласты клеток обкладки содержат ФС I, поэтому в них происходит только циклическое фотофосфорилирование; это означает, что в этих клетках не восстанавливается НАДФ+. Гранальные хлоропласты мезофильных клеток содержат обе фотосистемы, в них идет циклическое и нециклическое фотофосфорилирование с образованием АТФ и НАДФН.
Когда малат, образованный в мезофильных клетках, поступает в клетки обкладки, то при его декарбоксилировании происходит и окисление, и восстанавливается НАДФ+, необходимый для восстановления ФГК.
Таким образом, С4-цикл поставляет в цикл Кальвина и водород для восстановления СО2.
Эволюционно С3-цикл появился раньше, чем С4, он есть у водорослей. В древесных растениях цикла Хетча – Слэка нет. Это тоже подтверждает, что этот цикл возник позднее.
В заключение отметим, что, циклы Кальвина и Хетча – Слека действуют не изолированно, а строго координировано. Взаимосвязь этих двух циклов получила название «кооперативного» фотосинтеза.