

14.2.1.3. Скелет конечностей
У хордовых выделяются непарные и парные конечности. Непарные (спинные, хвостовой' и анальный плавники) являются основными органами передвижения у бесчерепных, рыб и в меньшей степени у хвостатых амфибий. У рыб возникают также парные конечности — грудные и брюшные плавники, на базе которых впоследствии развиваются парные конечности наземных четвероногих животных.
Подробнее рассмотрим происхождение и эволюцию парных конечностей.
У личинок рыб, а также у современных бесчерепных вдоль тела с обеих сторон тянутся боковые кожные складки, называемые метаплевральными (рис. 14.10). Они не имеют ни скелета, ни собственной мускулатуры, выполняя пассивную роль — стабилизацию положения тела и увеличение площади брюшной поверхности, облегчающие перемещение в водной среде. Вероятно, у предков рыб, переходящих к более активному образу жизни, в этих складках появились мышечные элементы и хрящевые лучи, связанные с сомитами по происхождению и поэтому расположенные метамерно. Такие складки, приобретя подвижность, могут выполнять роль рулей глубины, однако для изменения положения тела в пространстве большее значение имеют их передние и задние отделы, как наиболее удаленные от центра тяжести. Поэтому эволюция шла по пути интенсификации функций крайних отделов и ослабления функций центральных частей.
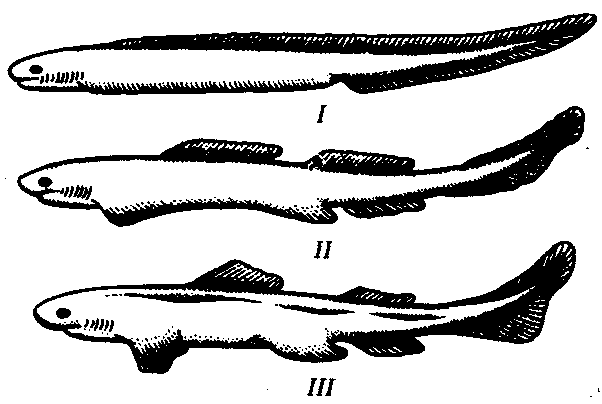
Рис. 14.10. Формирование передних и задних конечностей из метаплевральных складок: I—III—гипотетические этапы эволюции
В результате из передних отделов складок развились грудные, а из задних — брюшные плавники (рис. 14.10). Не исключено, что формированию только двух пар конечностей на боковых сторонах тела предшествовал распад сплошных складок на ряд парных плавников, большее значение из которых также имели передние и задние. Об этом свидетельствует существование ископаемых остатков древнейших низкоорганизованных рыб с многочисленными плавниками (рис. 14.11). За счет слияния оснований хрящевых лучей возникли плечевой и тазовый пояса. Остальные их участки дифференцировались в скелет свободных конечностей.

Рис. 14.11. Древняя акулообразная рыба с многочисленными парными конечностями
У большинства рыб в скелете парных плавников выделяют проксимальный отдел, состоящий из небольшого числа хрящевых или костных пластинок, и дистальный, в состав которого входит большое количество расположенных радиально сегментированных лучей. С поясами конечностей плавники соединены малоподвижно. Они не могут служить опорой телу при передвижении по дну или суше. У кистеперых рыб скелет парных конечностей имеет иное строение. Общее количество их костных элементов уменьшено, и они имеют более крупные размеры. Проксимальный отдел состоит только из одного крупного костного элемента, соответствующего плечевой или бедренной костям передних или задних конечностей. Далее следуют две более мелкие косточки, гомологичные локтевой и лучевой или большой и малой берцовым костям. На них опираются 7—12 радиально расположенных лучей. В соединении с поясами конечностей у такого плавника участвуют только гомологи плечевой или бедренной костей, поэтому плавники кистеперых рыб оказываются активно подвижными (рис. 14.12, А, Б) и могут использоваться не только для изменения направления движения в воде, но и для перемещения по твердому субстрату.
Жизнь этих рыб в мелких пересыхающих водоемах в девонском периоде способствовала отбору форм с более развитыми и подвижными конечностями. Наличие у них добавочных органов дыхания (см. разд. 14.3.4) стало второй предпосылкой выхода на сушу и возникновения других адаптации к наземному существованию, результатом чего явилось происхождение земноводных и всей группы Tetrapoda. Первые их представители — стегоцефалы — обладали семи- и пятипалыми конечностями, сохраняющими сходство с плавниками кистеперых рыб (рис. 14.12, B)

Рис. 14.12. Скелет конечности кистеперой рыбы (А), его основание (Б) и скелет передней лапы стегоцефала (В): I—плечевая кость, 2—локтевая кость, 3—лучевая кость
В скелете запястья сохранено правильное радиальное расположение костных элементов в 3—4 ряда, в пястье располагается 7—5 костей, а далее также радиально лежат фаланги 7—5 пальцев.
У современных земноводных количество пальцев в конечностях равно пяти или происходит их олигомеризация до четырех.
Дальнейшее прогрессивное преобразование конечностей выражается в увеличении степени подвижности соединений костей, в уменьшении количества костей в запястье сначала до трех рядов у амфибий и затем до двух — у пресмыкающихся и млекопитающих. Параллельно уменьшается также и количество фаланг пальцев. Характерно также удлинение проксимальных отделов конечности и укорочение дистальных.
Расположение конечностей в ходе эволюции также меняется. Если у рыб грудные плавники находятся на уровне первого позвонка и обращены в стороны, то у наземных позвоночных в результате усложнения ориентации в пространстве появляется шея и возникает подвижность головы, а у пресмыкающихся и особенно у млекопитающих в связи с приподнятием тела над землей передние конечности перемещаются кзади и ориентируются не горизонтально, а вертикально. То же касается и задних конечностей.
Многообразие условий обитания, предоставляемых наземным образом жизни, обеспечивает многообразие форм передвижения: прыжки, бег, ползание, полет, рытье, лазание по скалам и деревьям, а при возвращении в водную среду — и плавание. Поэтому у наземных позвоночных можно встретить как почти неограниченное многообразие конечностей, так и их полную вторичную редукцию, причем многие сходные адаптации конечностей в разнообразных средах многократно возникали конвергентно (рис. 14.13). Однако в процессе онтогенеза у большинства наземных позвоночных проявляются общие черты в развитии конечностей: закладка их зачатков в виде малодифференцированных складок, формирование в кисти и стопе вначале шести или семи зачатков пальцев, крайние из которых вскоре редуцируются и в дальнейшем развивается только пять (рис. 14.14).

Рис. 14.13. Скелет передней конечности наземных позвоночных. А—лягушка- Б—саламандра; В—крокодил; Г—летучая мышь; Д—человек: 1—плечевая кость, 2—лучевая кость, 3—кости запястья, 4—пясти, 5—фаланги пальцев, 6—локтевая кость
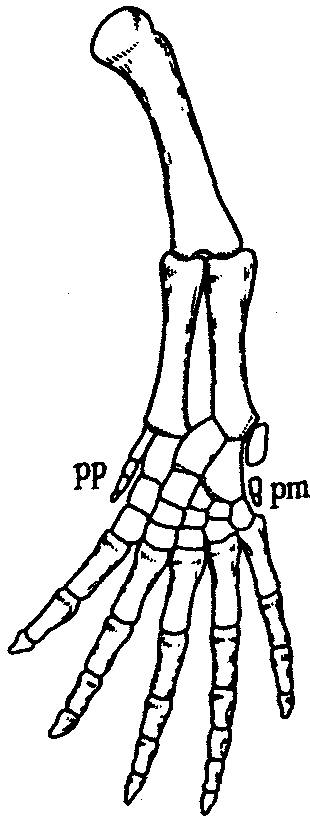
Рис. 14.14. Строение развивающейся конечности позвоночного: рр — prepollex, pin — postminimus — дополнительные рудиментарные I и VII пальцы
Интересно, что в эмбриогенезе высших позвоночных рекапитулирует не только строение конечностей предков, но и процесс их гетеротопии. Так, у человека верхние конечности закладываются на уровне 3—4-го шейных позвонков, а нижние — на уровне поясничных позвонков. В это же время конечности получают иннервацию из соответствующих отделов спинного мозга. Гетеротопия конечностей сопровождается формированием шейного, поясничного и крестцового нервных сплетений, нервы которых связаны с одной стороны с теми сегментами спинного мозга, из которых они вырастали в момент формирования конечностей, а с другой — с конечностями, переместившимися на новое место (рис. 14.15; см. также разд. 14.2.2.2).
В онтогенезе человека возможны многочисленные нарушения, ведущие к формированию врожденных пороков развития конечностей атавистического плана. Так, полидактилия, или увеличение количества пальцев, наследующаяся как аутосомно-доминантный признак, является результатом развития закладок дополнительных пальцев, характерных в норме для далеких предковых форм. Известен феномен полифалангии, характеризующийся увеличением числа фаланг обычно большого пальца кисти. В основе его возникновения — развитие трех фаланг в первом пальце, как это в норме наблюдается у пресмыкающихся и земноводных с недифференцированными пальцами конечностей. Двусторонняя полифалангия наследуется аутосомно-доминантно.
Серьезным пороком развития является нарушение гетеротопии пояса верхних конечностей из шейной области на уровень 1—2-го грудных позвонков. Эту аномалию называют болезнью Шпренгеля или врожденным высоким стоянием лопатки (рис. 14.16). Она выражается в том, что плечевой пояс с одной либо с двух сторон находится выше нормального положения на несколько сантиметров. В связи с тем что такое нарушение часто сопровождается аномалиями рёбер, грудного отдела позвоночника и деформацией лопаток, следует думать, что механизмы его возникновения — не только нарушение перемещения органов, но и обусловленное этим нарушение морфогенетических корреляций (см. § 13.4).
Сравнительно-анатомический обзор эволюции скелета хордовых свидетельствует о том, что скелет человека полностью гомологичен опорному аппарату предковых и родственных форм. Поэтому многие пороки его развития у человека можно объяснить родством млекопитающих с пресмыкающимися, земноводными и рыбами. Однако в процессе антропогенеза появились такие особенности скелета, которые характерны лишь для человека и связаны с его прямохождением и трудовой деятельностью. К ним относят: 1) изменения стопы переставшей выполнять хватательную функцию, выражающиеся в потере способности к противопоставлению большого пальца и появлению ее сводов, служащих для амортизации при ходьбе; 2) изменения позвоночного столба — его S-образный изгиб, обеспечивающий пластичность движений в вертикальном положении; 3) изменения черепа — резкое уменьшение его лицевой части и увеличение мозговой, смещение большого затылочного отверстия кпереди, увеличение сосцевидного отростка и сглаживание затылочного рельефа, к которому прикрепляются мышцы шеи и выйная связка; 4) специализация верхних (передних) конечностей как органа труда; 5) появление подбородочного выступа в связи с развитием членораздельной речи.
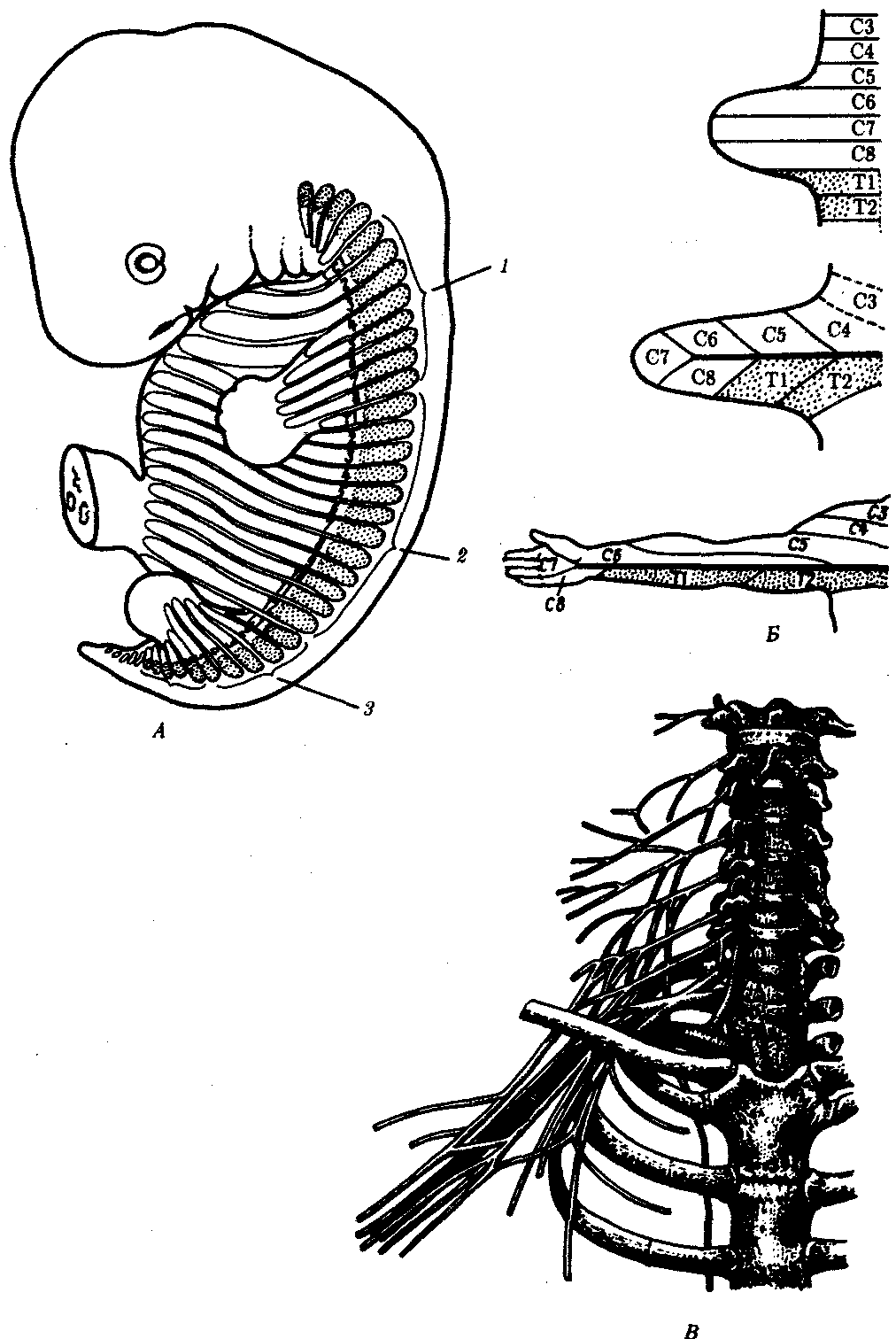
Рис. 14.15. Формирование передних конечностей, их гетеротопия и иннервация в онтогенезе человека. А—врастание шейных миотомов в формирующуюся переднею конечность зародыша; Б—развитие кожной иннервации руки; В— расположение шейного и плечевого сплетений, участвующих в иннервации руки:
1—шейные миотомы, 2—грудные миотомы, 3—поясничные миотомы; буквами С, Т, L обозначены шейные, грудные и поясничные сегменты

Рис. 14.16. Болезнь Шпренгеля (пояснение см. в тексте)
Несмотря на то что становление анатомо-морфологических особенностей скелета у человека, по-видимому, завершено, адаптации к прямохождению у него имеют, как и все адаптации вообще, относительный характер. Так, при большой физической нагрузке возможно смещение позвонков или межпозвонковых дисков. Человек, перейдя к прямохождению, утратил способность к быстрому бегу и передвигается значительно медленнее большинства четвероногих животных.
Естественно, что в ходе внутриутробного развития черты скелета, характеризующие человека как уникальный биологический вид, формируются на конечных его этапах либо даже, как, например, S-образная форма позвоночника, в раннем постнатальном периоде развития. Они являются фактически анаболиями, возникшими в ходе филогенеза приматов. Поэтому атавистические аномалии скелета, связанные с задержками развития признаков, характерных только для человека, встречаются наиболее часто. Они практически не снижают жизнеспособность, но дети, обладающие ими, нуждаются в ортопедической коррекции, гимнастике и массаже. К таким аномалиям относят легкие формы врожденного плоскостопия, косолапости, узкую грудную клетку, отсутствие подбородочного выступа и некоторые другие.