

Метаболізм вуглеводів. Анаеробне окислення глюкози
.docПлан
-
Загальні риси метаболізму вуглеводів
-
Ферментативні реакції анаеробного гліколізу
-
Регуляція гліколізу
-
Енергетика аеробного окислення глюкози
-
Використана література
1. Загальні риси метаболізму вуглеводів
Прості вуглеводи (моносахариди—глюкоза, фруктоза, галактоза, маноза) після всмоктування в кишковому тракті та проникнення всередину клітин підлягають метаболічним перетворенням, які становлять підґрунтя їх біоенергетичної функції. Безпосередній і найбільший внесок в утворення АТФ здійснює окислення глюкози до кінцевих продуктів катаболізму — діоксиду вуглецю та води. Крім того, певна кількість вуглеводів, що постійно надходять в організм з продуктами харчування, формують енергетичні депо метаболічного палива у вигляді глікогену та нейтральних жирів.
Основним моносахаридом, що в результаті катаболічних реакцій окислення до кінцевих простих продуктів утворює найбільшу кількість АТФ, є D-глюкоза, основним джерелом якої в організмі людини та тварин є крохмаль рослинних продуктів харчування. Інші прості вуглеводи, що надходять до організму в складі їжі (фруктоза — як компонент дисахариду сахарози, галактоза — як компонент лактози тощо), підлягають метаболічним перетворенням переважно після їх трансформації у фосфорні ефіри глюкози.
Основні шляхи внутрішньоклітинного катаболізму глюкози:
-
аеробне окислення, в результаті якого глюкоза розщеплюється до двоокису вуглецю та води;
-
гліколітичний шлях розщеплення (гліколіз), в результаті якого глюкоза утворює проміжні продукти катаболізму (піровиноградну або молочну кислоту).
Глюкоза, яка всмоктується в кров у кількості, що перевищує безпосередні енергетичні потреби організму, відкладається про запас у вигляді глікогену ("тваринного крохмалю") або використовується для синтезу триацилглщеролів жирової тканини.
Вторинні шляхи перетворення глюкози, що призводять до утворення біологічно важливих метаболітів:
-
пентозофосфатний шлях (шунт) окислення глюкози, в результаті якого утворюються необхідні для інших реакцій метаболізму фосфорні ефіри моносахаридів (пентоз, тріоз тощо) та відновлена форма НАДФ+ (НАДФН);
-
перетворення глюкози на глюкуронову кислоту;
-
перетворення глюкози на аскорбінову кислоту (метаболічний шлях функціонує лише в деяких видів тваринних організмів).
Гліколіз (шлях Ембдена-Мейергофа) — центральний шлях катаболізму глюкози, сукупність ферментативних реакцій, в результаті яких шостивуглецева молекула глюкози С6Н12О6 розщеплюється до двох тривуглецевих молекул піровиноградної або молочної кислоти. Гліколіз є шляхом катаболізму глюкози, в якому кисень не бере безпосередньої участі, проте, за рахунок наявності в гліколізі окислювально-відновлювальних реакцій, у результаті гліколітичного розщеплення глюкози генерується дві молекули АТФ.
Гліколіз є різновидом бродіння — біохімічного процесу, за рахунок якого забезпечують свою потребу в енергії у формі АТФ більшість існуючих на Землі анаеробних організмів або аеробів при функціонуванні в умовах недостатнього забезпечення молекулярним киснем. Поширеним типом бродіння в анаеробів є утворення з глюкози етилового спирту— процес, що каталізується ферментами дріжджів і широко використовується для виробництва алкогольних напоїв:
С6Н12О6 → 2 С2Н5ОН + 2 СО2
В організмі людини та тварин розрізняють:
- аеробний гліколіз, що супроводжується утворенням з однієї молекули глюкози двох молекул піровиноградної кислоти (пірувату):
С6Н12О6 → 2 С3Н4О3 (піруват);
Аеробний гліколіз можна також розглядати як проміжний (гліколітичний) етап аеробного окислення глюкози до кінцевих продуктів—двоокису водню та води;
- анаеробний гліколіз, що супроводжується утворенням з однієї молекули глюкози двох молекул молочної кислоти (лактату):
С6Н12О6 → 2 С3Н6О3 (лактат)
Для більшості тканин людини та вищих тварин в умовах нормальної життєдіяльності характерний аеробний гліколіз, тобто утворення з глюкози пірувату, який у подальшому окислюється до вуглекислого газу й води. Анаеробний гліколіз має місце переважно в м'язах при інтенсивній фізичній діяльності, тобто при відносній кисневій недостатності, та в деяких високоспеціалізованих клітинах (зокрема, в еритроцитах, в яких відсутні мітохондрії) або за певних патологічних умов (клітини злоякісних пухлин).
Реакції гліколізу перебігаютьу цитозолі клітини і каталізуються ферментами, що локалізовані в цьому компартменті.
Виділяють дві стадії гліколізу:
-
Розщеплення молекули глюкози до двох молекул фосфотріоз (гліцеральдегід-3-фосфату та діоксіацетонфосфату). Ця стадія включає в себе послідовність реакцій, які потребують витрати двох молекул АТР на кожну молекулу глюкози, що розщеплюється.
-
Перетворення двох молекул фосфотріоз на дві молекули пірувату (або лактату). Ця стадія включає в себе окислювально-відновлювальні реакції ("гліколі-тична оксидоредукція"), які супроводжуються генерацією чотирьох молекул АТР.
Таким чином, у результаті розщеплення однієї молекули глюкози в реакціях аеробного або анаеробного гліколізу сумарний вихід АТР складає дві молекули, що можна подати таким рівнянням:
D-глюкоза + 2 АДФ + 2 Фн → 2 піруват (лактат) + 2 АТФ
2. Ферментативні реакції анаеробного гліколізу
За умов анаеробного гліколізу (наприклад, в інтенсивно працюючих скелетних м'язах або в молочнокислих бактеріях) гліколі-тичний НАДН не віддає свої відновлювальні еквіваленти в дихальний ланцюг мітохондрій, а використовується для відновлення пірувату до L-лактату:
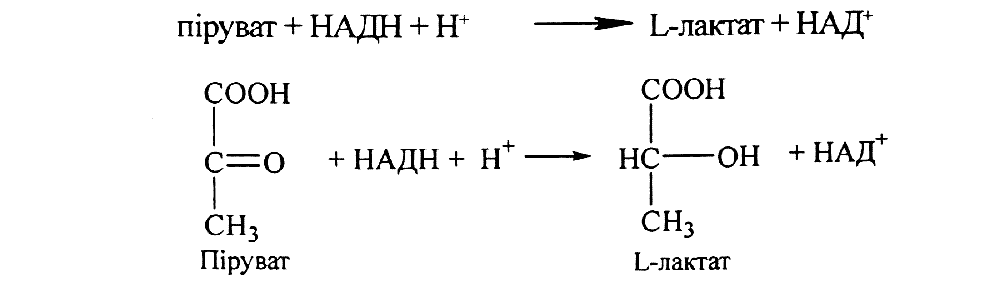
Реакція каталізується ферментом лактатдегідрогеназою, яка існує у вигляді п яти різних ізоферментних форм (ЛДГ1—ЛДГ5), що відрізняються за своїми кінетичними властивостями (КМ, Vмах, ступенем алостеричного інгібірування піруватом).
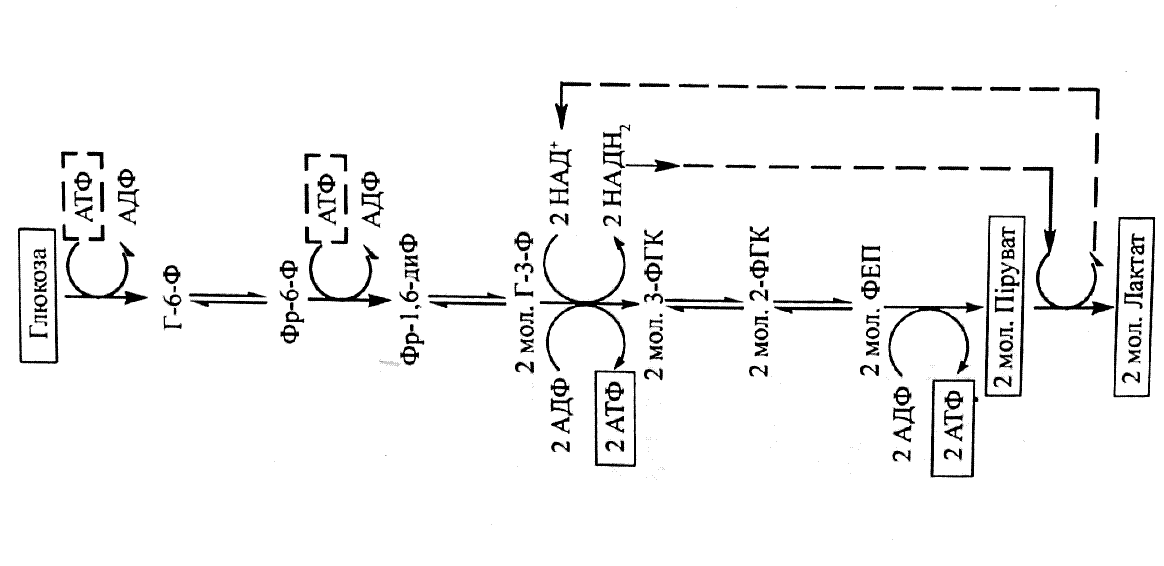
Мал. Загагьла послідовність ферментативних реакцій гліколізу
Таким чином, ферментативні реакції анаеробного гліколізу майже повністю співпадають із реакціями аеробного гліколізу, відрізняючись лише на етапі, що відбувається після утворення пірувату: при аеробному гліколізі піруват є субстратом перетворення на ацетилкоензим А та подальшого окислення, а при анаеробному гліколізі піруват відновлюється до лактату за рахунок НАДН, що утворився в реакціях гліколітичної оксидоредукції. Інакше кажучи, після утворення пірувату подальше його перетворення може відбуватися за одним із двох альтернативних шляхів, що залежать від стану окислювально-відновлювальних процесів у певній тканині:
- в аеробних умовах відбувається окисне декарбоксилювання пірувату доацетил-КоА, який у подальшому окислюється до СО, та Н,О в циклі Кребса; НАДН, що утворився при окисленні гліцеральдегід-3-фосфату, віддає свої відновлювальні еквіваленти на дихальний ланцюг мітохондрій через спеціальні човникові механізми;
- в анаеробнихумовах (або в умовах гіпоксії) реокислення гліколітичного НАДН відбувається за рахунок дії лактатдегідрогенази, яка відновлює піруват до лактату; течія лактат-дегідрогеназної реакції в даному напрямку генерує НАД+, що знову використовується для окислення гліцеральдегід-3-фосфату і подальшого накопичення лактату як продукту анаеробного гліколізу. Така послідовністьреакцій найбільш характерна для інтенсивно працюючихскелетних м'язів; крім скелетних м'язів та еритроцитів, клітини деяких інших органів та тканин (головного мозку, шлунково-кишкового тракту, мозкового шару нирок, сітківки та шкіри) частково задовольняють свої енергетичні потреби за рахунок анаеробного гліколізу, утворюючи молочну кислоту.
3. Регуляція гліколізу
Регуляція гліколізу здійснюється за рахунок впливу негативних та позитивних модуляторів (інгібіторів, активаторів) на каталітичну активність регуляторних ферментів, що каталізують незворотні реакції гліколізу:
-
гексокінази— для ферменту м'язів алостеричним інгібітором є продукт реакції глюкозо-6-фосфат;
-
фосфофруктокінази — інгібіторами є метаболіт трикарбонового циклуй цитрат та АТФ; активаторами — субстрат ферменту фруктозо-6-фосфат та АМФ; висока інтенсивність окислювальних процесів, що характеризується накопиченням у клітині субстратів ЦТК та АТФ, сприяє за рахунок даного механізму збереженню пулу глюкози; фосфофру ктокіназа є швидкість-лімітуючою реакцією гліколізу;
-
піруваткінази — фермент інгібується АТФ, а також субстратами циклу лимонної кислоти — ацетил-КоА та жирними кислотами, що забезпечує гальмування гліколізу в умовах високої інтенсивності окислювальних процесів; печінкова ізоформа піруваткінази регулюється за допомогою ковалентної модифікації — цАМФ-залежного фосфорилювання (дефосфорильована форма активна, фосфорильована — неактивна); крім того, піруваткіназа гепатоцитів є Індукованим ферментом, синтез якого стимулюється в умовах підвищеного надходження з їжею вуглеводів та зростання рівня інсуліну.
Гальмування реакцій гліколізу за рахунок пригнічення в умовах активного клітинного дихання каталітичних активностей фосфофруктокінази та піруваткінази є молекулярною основою ефекту Пастера.
4. Енергетика аеробного окислення глюкози
У процесі анаеробного окислення глюкози витрачається 2 молекули АТФ (при фосфорилюванні глюкози з утворенням глюкозо-6-фосфату і при перетворенні фруктозо-6-фосфату на фруктозо-1,6-дифосфат), а синтезується 4 АТФ (по дві у реакціях гліцеральдегід-3-фосфат → 3-фосфогліцерат та фосфоенолпіруват → піруват).
Таким чином, різниця у затраченій і утвореній кількості АТФ складає 2 молекули.
У зв'язку зі значно більшою енергетичною ефективністю аеробного окислення глюкози, порівняно з гліколізом, останній процес розглядається як еволюційно більш прадавній шлях катаболізму глюкози, що мав першорядне значення в умовах відсутності в первісній земній атмосфері кисню.
5. Використана література
-
«Биологическая химия» изд. «Медицина» 2000г. Е.С. Северин, Т.Л. Алейникова, Е.В. Осипов
-
Гунський Ю.І. – Біологічна хімія: Підручник. –Київ-Тернопіль: Укрмедкнига, 2000