

ОБ
ОРГАНИЗАЦИИ ФУНКЦИЙ СЛУХОВОЙ СИСТЕМЫ
Рассмотрение
вопроса об организации функции слуховой
системы неразрывно связано с представлением
о той задаче, которую выполняет эта
система.
Выше
(см. главу «Общая характеристика слуха
позвоночных») задача, выполняемая
слуховой системой, определялась как
установление свойств источников
звукоиз л учения, и в первую очередь
источников излучения сложных звуков
биологического происхождения. Однако
благодаря недостаточности наших знаний
вопрос об организации функции слуховой
системы может быть рассмотрен лишь с
точки зрения более узкой задачи,
выполняемой этой системой: восприятия
определенных параметров звукового
сигнала или некоторой их совокупности.
ЧАСТОТНОЕ
РАЗЛИЧЕНИЕ
Развитие
представлений о функции слуховой
системы исторически оказалось наиболее
тесно связанным с представлениями
о механизмах восприятия одного из
параметров звука, а именно частоты
колебаний. Эта связь не была случайной,
она вытекала, во-первых, из знаний
отношений между частотой колебания
струн и ощущением высоты тонов и
музыкальных интервалов, во-вторых,
из физических и математических
представлений об анализе колебательных
явлений. Обобщение этих знаний в труде
Гельмгольца «Учение о тональных
ощущениях, как физиологическая
основа теории музыки» (Helmholtz, 1863) являлось
первым систематическим изложением
свойств слуха и представлений о
механизмах анализа частоты звуковых
колебаний, осуществляемого ухом. В
этих представлениях Гельмгольц
основывался на следующих положениях.
1.
Любая периодическая форма звуковых
колебаний может быть составлена,
согласно теореме Фурье, из суммы простых
(синусоидальных) колебаний, обладающих
частотами, кратными основной частоте
этих колебаний.
286Глава 10
S. Ухо
осуществляет разложение любого
периодического звукового колебания
на отдельные простые составляющие,
которые вызывают ощущение тона
определенной высоты (правила Ома).
3.
Так как физически разложение периодического
звукового колебания любой формы на
отдельные составляющие может быть
осуществлено набором резонаторов, то
очевидно, что в структурах улитки должны
существовать элементы, обладающие
подобными свойствами.
Вначале
Гельмгольц приписывал свойства
резонирующих элементов «Кортневым
дугам», однако в связи с появлением
новых данных о гистологическом строении
улитки Гельмгольц пришел к заключению,
что базилярная мембрана, которая
постепенно расширяется от основания
к вершине, благодаря большому поперечному
натяжению может рассматриваться как
упорядоченно расположенный набор
механических резонаторов, настроенных
на частоты, воспринимаемые ухом.
Этот
набор механических резонаторов разлагает
воздействующие на ухо периодические
колебания любой формы на простые
составляющие: каждый резонирующий
элемент раздражает определенные,
связанные с ним волокна слухового
нерва; возбуждение этих волокон вызывает
ощущение соответственной высоты тона,
являющегося простейшим элементом
музыкального восприятия. Предел
различения одного тона от другого
должен определяться частотной
избирательностью резонаторов (см. главу
«Психоакустика», стр. 328).1
Упорядоченное
расположение настроенных на разные
частоты резонаторов по длиннику улитки
определяет передвижение максимума
колебаний от основания до вершины
улитки при изменении частоты воздействующих
звуковых колебаний от высоких к низким.
Подобное представление получило в
дальнейшем обозначение пространственного
принципа анализа частоты звука.
Как
известно из предыдущего изложения (см.
главу «Механизм и физиология
наружного, среднего и внутреннего
уха»), данные прямого измерения колебаний
базилярной мембраны улитки не согласуются
с представлением Гельмгольца о
базилярной мембране как наборе
резонирующих элементов. Частотная
избирательность базилярной мембраны
оказывается значительно меньшей, чем
предполагал Гельмгольц. Однако сам
принцип пространственной проекции
частот, разработанный Гельмгольцем,
сохраняет свое полное значение в
настоящее время (Fletcher, 1929; Bekesy, 1960).
1
Одной из характеристик частотной
избирательности является затухание
амплитуды относительно максимума,
наблюдаемого на частоте резонанса, при
сдвиге частоты на одну октаву. Из
допущенной Гельмгольцем зависимости
амплитуды колебаний резонаторов уха
от частоты вытекает, что частотная
избирательность этих резонаторов могла
бы быть охарактеризована величиной
31.4 дб на октаву.
287
Уже
во времена Гельмгольца и Ома и особенно
в последующее время обращалось внимание
на то, что для механизма восприятия
высоты тонов может иметь значение не
только анализ частоты колебаний,
осуществляемый набором резонаторов
или фильтров, но и анализ периодичности
звукового сигнала (Schouten, 1940; Schouten et al.,
1962).
Несмотря
на то что частота (/) и период (Т)
гармонического колебания —- обратные
величины методы их физического
анализа
неоднозначны (Licklider, 1951).
Если
частота измеряется местом возникновения
максимальной амплитуды колебания, то
периодичность измеряется длительностью
интервалов между максимумами (или
минимумами) амплитуд колебаний.
Необходимость
поисков физиологического механизма
измерения временных интервалов
диктуется целым рядом наблюдений
по восприятию высоты сложных звуков,
которые трудно объяснимы с точки
зрения принципа пространственного
анализа частоты (Schouten, 1940; Schouten et al.,
1962; дискуссия в кн.: Bekesy, 1963; см. также
главу «Психоакустика»).
Электрофизиологические
данные, которые были приведены в в главе
«Электрические проявления деятельности
путей и центров слуховой системы»,
свидетельствуют о том, что в разрядах
волокон слухового нерва кошки и обезьяны
воспроизводятся длительности
периодов или кратные им отношения
воздействующей на ухо звуковой частоты
вплоть до 4—5 кгц (рис. 73) (Kiang, 1965а; Rose et
al., 1967).
При
действии щелчков (Kiang, 1965а; рис. 103), а
также широкополосных шумов в группах
волокон, характеристичная частота
которых не выше 4.0 кгц, наблюдаются
периодические разряды с периодом,
соответствующим характеристичной
частоте (de Boer, 1968). При исследовании
суммарных ответов в слуховом нерве и
кохлеарных ядрах наблюдается синхронизация
с частотой раздражения вплоть до
3—5 кгц (Davis, 1935; Kemp et al., 1937; Гер- шуни,
1940).
Все
эти данные не оставляют сомнения в том,
что в разрядах волокон слухового нерва
содержатся сведения о длительности
периода.
Центральные
механизмы, которые могли бы обрабатывать
эти сведения, в настоящее время совершенно
не ясны.
Оба
механизма передачи сведений: о частоте
по месту раздражения и о периодичности
по интервалам между разрядами импульсов
отнюдь не исключают, а дополняют друг
друга. Одновременное использование
обоих механизмов должно иметь место
при восприятии звуков в низкочастотном
диапазоне от 50 до 4000— 4500 гц, а также
сложных звуков, щелчков и шумов
низкочастотными группами элементов.
Необходимо отметить, что диапазон
288
частот,
характерный для звуков речи и музыки,
укладывается в предел, для которого
действительны оба механизма.
Имеются
данные, позволяющие предполагать, что
в процессе эволюции у таких наземных
млекопитающих, как хищники (например,
кошка), приматы и человек произошло
возрастание чувствительности в
области частот ниже 4000 гц (Masterton et al.,
1969), т. е. в области частот, в которой
могут осуществлять свое действие оба
механизма, в отличие от области более
высоких частот, включая ультразвуковые,
при восприятии которых сведения о
периодичности сигнала, как можно судить
по имеющимся данным, должны
отсутствовать.
Для
разбираемого вопроса существенны
данные прямого электрического раздражения
у человека слуховых нервных волокон,
идущих от разных участков улитки (см.
главу «Слуховое восприятие у человека
в условиях электрического раздражения
разных отделов слуховой системы»). Эти
данные свидетельствуют о зависимости
ощущения высоты от двух факторов: а)
пространственного расположения
раздражаемых нервных волокон в улитке,
б) частоты следования раздражающих
электрических стимулов. Эти факты
служат сильным аргументом в пользу
существования двух отличных
нейрофизиологических механизмов,
определяющих ощущение высоты.
Все
изложенное позволяет полагать, что в
диапазоне более высоких звуковых частот
(выше 4—5 кгц) частотное различение
может обеспечиваться только механизмом
проекции по месту раздражения, а в
диапазоне очень низких частот (ниже
30—50 гц) — механизмом воспроизведения
периодичности сигнала. В области
средних частот различение, по-видимому,
обеспечивается при участии обоих
механизмов.
Как
можно судить по данным исследования
частотно-различительных свойств
улитки, указанные пределы, особенно в
области низких частот, обнаруживают
существенные отличия у разных
млекопитающих (Bekesy, 1944). Следует, однако,
указать, что пределы воспроизведения
центральными отделами слуховой системы
периодичности сигнала, а также центральная
проекция частот изучены в настоящее
время лишь у немногих представителей
млекопитающих (наиболее систематически
у кошки).
В
предыдущем изложении (см. главу «Механика
и физиология наружного, среднего и
внутреннего уха») при рассмотрении
данных об амплитуде колебаний
базилярной мембраны при действии звуков
различных частот уже обращалось внимание
на то, что частотная избирательность2
этого отдела улитки ниже частот-
2
Количественно частотная избирательность
может быть охарактеризована, как
уже говорилось, степенью затухания в
децибелах на октаву или коэффициентом
Q,
выражающим отношение между оптимальной
частотой (/) и шириной полосы (Д/) при
данном уровне интенсивности (см. главу
19
Сенсорные системы 289
йой
избирательности волокон слухового
нерва, устанавливаемой по частотнопороговым
кривым.
Следует
указать, что значение частотнопороговых
кривых как метода суждения о частотной
избирательности отдельных эле-^ ментов
слуховой системы должно рассматриваться
с определенными ограничениями. Как
показывает само обозначение, построение
частотнопороговых кривых основывается
на использовании реакций, возникающих
на пороговые для данного элемента
интенсивности звуков различных
частот. Для суждения о частотной
избирательности нервного элемента
слуховой системы, помимо порогового
метода, используется метод равных
импульсных ответов; в этом случае
критерием служит ответ, выражаемый
определенным числом нервных импульсов.
Этот
более общий метод определения частотной
избирательности по контурам равных
импульсных ответов дает возможность
устанавливать зависимость ответа от
частоты звука на всем диапазоне
интенсивностей, на который отвечает
данная нервная единица. Частотнопороговая
кривая является по существу частным
случаем измерения, в котором за
равноимпульсный контур принимается
пороговый ответ единицы.
Определение
частотной зависимости для волокон
слухового нерва (Sachs, Kiang, 1968) и нейронов
кохлеарных ядер (Greenwood, Maruyama, 1965) при
разных значениях равноимпульсных
ответов (от 30 до 200 имп./ сек. для волокон
слухового нерва) показывает, что острота
частотно-равноимпульсных кривых
закономерно возрастает при
соответственном возрастании уровня
интенсивности звука. Это явление
особенно выражено для волокон
слухового нерва.
Таким
образом, частотная избирательность
ответа отдельной единицы возрастает
(обостряется) при увеличении интенсивности
раздражения до определенного уровня
(рис. 101).
При
оценке как частотнопороговых, так и
частотно-равноимпульсных кривых,
по которым судят о частотно-избирательных
свойствах слуховых нервных единиц,
возникает общий вопрос о возможных
механизмах возрастания (обострения)
частотной избирательности, выражаемой
в ограничении области частот, на которые
отвечает нервная единица.
При
рассмотрении этого вопроса следует
прежде всего подчеркнуть особенности
оценки функции слуховой системы по
реакциям отдельного ее элемента.
Каждый отдельный элемент слуховой
системы, будь то волокно спирального
ганглия или нейрон вышележащего
отдела, связан с определенными участками
поверхности улитки, на котором расположено
множество рецепторных элемен-
«Психоакустика»,
стр. 308). Для частотнопороговых кривых
обычно в физиологических исследованиях
принимается уровень интенсивности,
равный 10 дб над порогом оптимальной
частоты.
290
тов,
возбуждаемых при действии звуков разных
частот, используе-
мых при определении
частотно-избирательных свойств единицы.
Какое
значение должно иметь возбуждение
других рецептор-
ных
и нервных единиц слуховой системы при
определении ча-
стотно-избирательных
свойств данной единицы?
Это
общий вопрос об организации связей в
рецептивном поле
нервной
единицы, который изучался для всех
сенсорных систем,
и особенно подробно
для зрительной системы (см. часть I
тома
«Физиология сенсорных систем»).
Рассмотрим
с точки зрения ор-
ганизации
рецептивного поля оп-
ределение
частотнопороговой кри-
вой. Нахождение
частоты, поро-
Рис.
101. Частотные зоны ответов нейронов
слухового нерва (А) и кохлеар-
ного
ядра (Б), полученные при подсчете равного
количества импульсов
на каждой
частоте (А
— по: Sachs, Kiang, 1968; Б
—
по: Greenwood, Maru-
yama, 1965).
На
А:
по оси абсцисс —
частота звука в кгц, по
оси ординат слева —
смещение стремечка в мкм, справа
—
интенсивность звука в дб. Цифры
над кривыми —
частота (имп./сек.). На В:
по оси абсцисс —
частота звука в кгц (верхний
ряд цифр)
и место расположения волосковых клеток
на базилярной мембране, которые
раздражаются соответствующей частотой,
в мм от верхушки улитки (нижний
ряд цифр)', по оси ординат —
уровень звукового давления в дб.
Нижняя
кривая —
50 имп. в 1 сек., верхняя
—
240 имп. в 1 сек.
говая
интенсивность для которой наиболее
низка (характеристичная или оптимальная
частота), может рассматриваться как
способ установления преимущественной
связи данного элемента с определенным
участком рецепторной поверхности
улитки. При низком абсолютном значении
порога на оптимальной частоте, что
характерно для многих элементов,
раздражаемая область должна быть
наименьшей и одновременно с исследуемым
элементом должно возбуждаться
относительно небольшое количество
единиц с близкими оптимальными
частотами. Чем больше повышается порог
исследуемого элемента при сдвиге
частоты от оптимальной (т. е. чем острее
его частотнопороговая кривая), тем
больше должна быть интенсивность
раздражения тех неисследуемых эле
19*
291

ментов,
оптимальная частота которых соответствует
сдвигу частоты на частотнопороговой
кривой.
Естественно
предположение по аналогии с другими
сенсорными системами, что одновременное
возбуждение элементов, обладающих
разными оптимальными частотами, и,
следовательно, расположенных в
разных участках улитки, входящих, однако
в рецептивное поле исследуемого
элемента, вызывает тормозное
взаимодействие, определяющее
ограничение полосы воспринимаемых
частот, т. е. явления обострения частотной
избирательности нервных элементов
по сравнению с частотной избирательностью
базилярной мембраны.
Предположение
о тормозном взаимодействии как
нейрофизиологическом механизме
разнообразных явлений обострения
ответа слуховой системы на стимул
многократно обсуждалось в литературе
(Bekesy, 1967; см. также главу «Психоакустика»).
Наиболее
прямой путь экспериментального изучения
этого взаимодействия выражается,
во-первых, в исследовании результатов
действия двух пространственно разнесенных
на рецепторной поверхности раздражения,
как это впервые в наиболее ясной форме
было показано на рецепторной поверхности
глаза (Hartline et al., 1961); во-вторых, в
одновременном исследовании двух
пространственно разнесенных нервных
элементов. К сожалению, последнего рода
исследования до сих пор еще не были
осуществлены в слуховой системе.
В
слуховой системе экспериментальное
изучение тормозного взаимодействия
осуществляется в условиях одновременного
действия двух отличных по частоте
звуков, которые проецируются на различные
участки рецепторной поверхности улитки.
При
действии звука, состоящего из двух
компонентов, отношение обоих
компонентов может видоизменяться по
ряду признаков: частоте, уровням
интенсивности, временным интервалам.
Частота одного из используемых
компонентов подбирается в соответствии
с оптимальной частотой исследуемой
нервной единицы.
Явление,
выражающее взаимодействие элементов
системы при раздражении разными
компонентами сигнала, для всех волокон
слухового нерва носит однообразный
характер, что дает’возможность
изобразить его в условном (идеализированном)
виде (рис. 102).
Приведенные
данные о действии двух тонов несомненно
свидетельствуют, что обострение
частотной избирательности может
определяться развитием тормозных
явлений.
Поэтому
общая логическая схема механизма
обострения может быть изображена в
виде схемы тормозных связей соседних
элементов друг с другом, как это
представлено в ряде работ (Furman, Frishkopf,
1964; Молчанов, Лабутин, 1966; Любинский,
Позин, 1967). Однако тормозное взаимодействие,
наблюдаемое в^волок- нах слухового
нерва, представляет значительные
трудности для
292
толкования
его механизмов. Прежде всего в улитке
неясны связи
между нервными волокнами
или клетками спирального ганглия,
которые
могли бы определять развитие афферентного
торможе-
ния. Поэтому для объяснения
явлений обострения на уровне
волокон
слухового нерва, а также наличия
тормозных зон вво-
дятся дополнительные
предположения о распространении
ге-
нераторных потенциалов по
дендритам (Любинский, Шараев,
1968) или
вызванных ими токов по жидким средам
улитки (Fur-
man, Frishkopf, 1964).
При
оценке возможных механизмов обострения
частотной из-
бирательности
в волокнах слухового не
что
частотно-амплитудные зависимости
для
механических структур улитки из-
вестны
пока только для базилярной мем-
браны,
а не для других механических
структур,
которые непосредственно свя^
заны
с возбуждением волосковых кле-
ток.
Поэтому возможно, что более
высокая
избирательность волокон слу-
Рис.
102. Тормозные и возбуждающие
частотные
зоны ответов волокон слухового
нерва
(по: Sachs, Kiang, 1968). 0
По
оси абсцисс —
частота звука; по
оси ординат —
уровень интенсивности. f0
—
опти-
мальная частота. 1
— треугольником
показан средний уровень интенсивности,
при ко-
тором проводилось измерение
тормозных зон; 2
—
частотная возбуждающая зона ответа;
3
—
тормозные зоны (заштрихованы').
хового
нерва определяется в определенной
степени механическими преобразованиями
в звеньях: базилярная мембрана — волоски
рецепторных клеток.
Сопоставление
частотнопороговых кривых и тормозных
зон в разных отделах позволяет отметить
характерные отличия между явлениями,
наблюдаемыми на уровне волокон слухового
нерва и вышележащими отделами слуховой
системы.
Хотя
в слуховом нерве описывается разделение
входящих в него волокон на группы по
некоторым свойствам, в первую очередь
по значениям порогов, в целом для волокон
слухового нерва обнаруживаются некоторые
общие характеристики, типичные для
всей популяции волокон. Это дает
возможность описать свойства
некоторого типичного (усредненного)
нервного волокна. К этим свойствам
относятся, в частности, характеристики
возбуждающих и тормозных областей;
воспроизведение во временных
интервалах между нервными разрядами
периодичности низких звуковых частот
(до 4500—5000 гц); возрастание остроты
частотно-пороговых кривых при
увеличении оптимальной частоты
293
волокон
и др. (см. раздел «Слуховой нерв» главы
«Электрические* проявления деятельности
слуховой системы»).
На
уровне кохлеарных ядер, оливарного
комплекса, особенно^ заднего двухолмия,
появляются нейроны с отличными
характеристиками, различными по
остроте областями ответа и типами
разрядов, симметричными и несимметричными
тормозными зонами' с неоднозначной
зависимостью частотно-избирательных
свойств* от длительности сигнала для
разных типов нейронов. В этих центральных
отделах становится уже невозможным
описать свойства’ некоторого единого
усредненного нервного элемента в
отличие от* нервных волокон, составляющих
слуховой нерв у таких млекопитающих,
как кошка.
Возрастающее
разнообразие типов ответов нейронов
на сигнал свидетельствует о появлении
иных свойств функционально-структурной
организации центральных отделов. Эти
свойства не могут рассматриваться
изолированно лишь для случая анализа
частоты стационарного звука.
ВРЕМЕННЫЕ
ХАРАКТЕРИСТИКИ СИГНАЛА
И
ИХ ВЫДЕЛЕНИЕ
Всякий
реально существующий звук обладает
характеристиками, которые не являются
стационарными и изменяются во
времени. Это относится прежде всего к
началу и концу сигнала. Момент включения
и выключения звукового сигнала
характеризуется изменением амплитуды
в последовательные отрезки времени.
Эти
изменения амплитуды могут в разных
случаях протекать, с различной скоростью;
происходящие изменения амплитуды во>
времени связаны определенным образом
со спектральной характеристикой
сигнала (Харкевич, 1962).
При
прямоугольной форме сигнала изменение
амплитуды является теоретически
мгновенным, а мгновенный спектр в эти
моменты бесконечно широким.3
При постепенном нарастании амплитуды
мгновенный спектр окажется соответственно
более узким.
Подобные
переходные или нестационарные явления
будут характеризовать начальный и
конечный отрезки, за которыми при
достаточной длительности сигнала будет
следовать установившийся (стационарный)
участок, на протяжении которого сигнал
сохраняет постоянные характеристики.
Для
очень коротких сигналов, подобных
щелчку, стационарный участок может
отсутствовать.
3
Практически любая акустическая система
имеет временные задержки и спектральные
ограничения и поэтому воспроизводит
идеализированные свойства сигнала с
той или иной степенью искажения.
294
Кроме
переходных явлений включения и
выключения, сигналы могут характеризоваться
различными видами модуляций (амплитудной,
частотной, фазовой). Амплитудная и
частотная модуляции являются характерными
для естественных биологических
сигналов.
Изменение
амплитуды, частоты и фазы находит
типичные выражения в форме и спектре
сигнала (Харкевич, 1962).
При
электрофизиологическом исследовании
зависимость ответа от формы сигнала
выступает весьма резко в картине
возникающих суммарных ответов.
Предельно крутой, короткий и широкополосный
сигнал, каковым является щелчок, может
служить примером наиболее эффективного
раздражения для вызова суммарных
ответных реакций во всех отделах
слуховой системы (рис. 103).
Уменьшение
крутизны фронта сигнала вызывает
падение величины ответа вплоть до
полного его исчезновения (рис. 104, 105).
Механизм этого явления наиболее ясно
выступает при рассмотрении суммарного
ответа волокон слухового нерва.
Суммарный
ответ слухового нерва, измеряемый по
наиболее выраженному начальному
компоненту Нг,
может рассматриваться как результат
сложения импульсов, возникающих в
слуховых нервных волокнах. Амплитуда
суммарного ответа, зависящая от числа
участвующих в сложении нервных
импульсов,4
определяется не только числом возбуждаемых
волокон, но и совпадением импульсов
во времени (синхронизацией), зависящим
от степени совпадения латентных
периодов.
Сравнение
латентных периодов группы волокон,
обладающих разной характеристичной
частотой, достаточно ясно свидетельствует
о значении этого фактора (рис. 103). Так
суммарный ответ определяется сложением
разрядов только коротколатентных
волокон с высокой характеристичной
частотой, идущих от базального
завитка улитки (см. на рис. 103 волокона
3 и 4).
Возрастание
амплитуды суммарного ответа Нг
при возрастании интенсивности щелчка
свидетельствует о наличии в популяции
нервных волокон, участвующих в образовании
суммарного ответа, волокон, сильно
отличающихся по значению порогов (см.
раздел «Слуховой нерв» главы 7, а также
Davis, 1961).
Уменьшение
крутизны фронта сигнала (на рис. 104
широкополосного шума) вызывает
уменьшение и исчезновение суммарного
ответа. Это явление происходит не за
счет прекращения ответа отдельных
волокон, а за счет возрастания разброса
латентных периодов этих ответов в
совокупности волокон, обладающих
разными порогами (рис. 104).
4
О предполагаемой физической модели
сложения электрических напряжений
в совокупности параллельно расположенных
нервных волокон см.: Stevens, Davis, 1938.
295
256
Рис.
103. Соотношение разрядов волокон
слухового нерва и суммарного
потенциала действия, регистрируемого
от круглого окна улитки (по: Kiang, 1965а).
Вверху
—
суммарный потенциал действия, внизу
— постстимульные
гистограммы 4 нейронов (1—4)
на
звуковые щелчки.
На
постстимульных гистограммах по
оси абсцисс — время
в мсек.; по
оси ординат —
количество импульсов, 0.64;
2.0; 5.3; 9.0 —
значения оптимальной частоты нейронов
в кгц. Для нейронов I, 2
интервалы между пиками на гистограммах
соответствуют периоду характеристичных
частот (0.64
и
2
кгц).
128
64
О
128


Таким
образом, суммарный ответ на очень
короткий сигнал или длительный, но
круто нарастающий сигнал подчеркивает,
благодаря явлению синхронизации,
реакцию на крутизну фронта нарастания
сигнала. Поэтому начальный синхронизированный
ответ
для короткого и длин-
ного сигнала
мало отлича-
ются друг от друга; в
по-
следнем случае ответ явля-
ется
одновременно выраже-
нием реакции
на начало
сигнала (рис. 105).
Подобного
рода явление
имеет место во всех
отделах
слуховой системы. Однако
для
центральных отделов си-
стематически
растет постоян-
ная времени нарастания
амп-
литуды сигнала, при которой
может
наблюдаться началь-
ный синхронизированный
от-
вет. Так, в слуховой коре
начальная
позитивная волна
первичного ответа
наблюда-
ется и при очень
медленно
нарастающих сигналах
(по-
стоянная времени более
Рис.
104. Суммарный нервный от-
вет улитки
(Л) и первичный ответ
слуховой зоны
коры (Б)
при раз-
личной постоянной времени
зву-
кового сигнала (Л — по:
Забоева,
1966; Б
—
по: Гершуни, 1965).
Цифры
слева —
величина постоянной времени в мсек.; к
—
контрольные осциллограммы, снятые в
отсутствии звукового раздражения.
Наверху
—
осциллографическая запись звукового
сигнала (белого шума); внизу
—
суммарные нервные ответы улитки (А)
и
первичные ответы слуховой коры (Б).
100
мсек.). Таким образом, начальный суммарный
синхронизированный ответ в коре
обнаруживается при полном нарушении
синхронизации в нижележащих уровнях,
и в первую очередь в слуховом нерве-
(рис. 104, 5).
Изложенные
данные, основанные на использовании
суммарных ответов, показывают зависимость
ответов слуховой системы от постоянной
времени сигнала и значительные отличия
в характеристиках этой зависимости
в волокнах слухового нерва по сравнению
с вышележащими отделами, особенно
слуховой областью коры.

Исследование
реакций отдельных нервных единиц
слуховой системы на изменения постоянной
времени нарастания и спадения сигнала
осуществлены для нейронов кохлеарных
ядер и задних холмов. Результаты
измерений обнаруживают резко отличные
реакции нейронов разных типов на
изменения этого временного параметра
сигнала. Так, коротко латентные,
обладающие малой временной суммацией,
фазические нейроны реагируют резким
повышением порогов на возрастание
постоянной времени сигнала; при этом
частотная избирательность, как можно
судить по остроте частотнопороговых
кривых, не претерпевает заметных
изменений
А Б
Рис.
105. Суммарные нервные ответы улитки на
сигналы разной длительности (по: Гершуни,
1965).
А
— длительность звука 0.8 мсек., Б
—
20 мсек. Наверху
—
осцил-
лографическая запись звукового
сигнала (белого шума); внизу
—
суммарный
нервный ответ улитки.
(рис.
87, Б,
2).
Длиннолатентные нейроны, обладающие
выраженной временной суммацией
(тонические и пачковые), не реагируют
заметным изменением порогов даже
при очень значительном возрастании
постоянной времени сигнала; частотная
избирательность нейрона в этих условиях
обостряется (рис. 87, Б,
1).
Таким
образом, при изменении крутизны фронта
сигнала должно происходить перераспределение
возбуждаемой популяции нервных единиц
вышележащих отделов слуховой системы,
отличающихся по своим временным
характеристикам.
В
условиях действия тональных сигналов
(в отличие от широкополосных шумов)
переходные явления характеризуются
особенно выраженными относительными
изменениями спектра. Перераспределение
возбуждаемых нервных единиц, обладающих
временными характеристиками,
выделяющими разные моменты действия
сигнала и по-разному изменяющими свою
частотную избирательность в
зависимости от временных свойств
сигнала,, оказывается особенно
существенным для получения одновременных
сведений о временных и спектральных
признаках переходные явлений^
298
Помимо
выраженных реакций на переходные
явления, следует’ обратить внимание
на наличие в центральных отделах
слуховой системы (задних холмах и
слуховой коре) нервных единиц, ответ
которых резко уменьшается при возрастании
длительности сигнала. Так, на рис.. 106
представлен нейрон заднего двухолмия
белой крысы, разряды которого для трех
шумовых сигналов длительностью в
2, 5 и 50 мсек., обнаруживают резкое падение
импульсации при возрастании
длительности сигнала.. Это явление
совершенно отлично от явлений
адаптации,, которые наблюдаются на
уровне волокон слухового нерва (см.
раздел «Слуховой нерв» главы 7).
На
уровне слухового нерва, во всяком случае
у такого систематически изученного
животного, как кошка, не описана
дифференциация волокон на группы,
обладающие различными реакциями на
временные признаки сигнала.
Таким
образом, появление групп нервных
элементов, по-разному реагирующих
на временные признаки сигнала, может
рассматриваться как характерная
особенность функций центральных отделов
слуховой системы млекопитающих (кошка,
белая крыса). Следует указать, что у
других позвоночных, в частности
земноводных, слуховая система которых
начинает подвергаться систематическому
изучению в последнее время (Frishkopf,
Goldstein, 1963; Бибиков, 1969, 1970), относительное
значение разных отделов может быть
существенно отличным от описанного
для млекопитающих.
ВОСПРИЯТИЕ
СЛОЖНЫХ ЗВУКОВ
Под
сложными звуками понимают обычно звуки,
обладающие разными компонентами, как
независимыми, так и взаимосвязанными.
Изменение отдельных компонентов
сложного звука, а также получение
определенных сочетаний этих компонентов
является распространенным приемом
экспериментального исследования.
Одним из широко используемых в
эксперименте сложных звуков является
амплитудномодулированный звук.
Исследование слуховой функции при
помощи амплитудномодулиро- ванных
сигналов интересно потому, что дает
возможность относительно независимо
осуществлять изменение временных
признаков сигнала путем изменения
частоты модуляции (g) и спектральных
свойств путем изменения частоты
модулируемого сигнала (/); спектр такого
сигнала будет выражаться тремя
компонентами (f*
f—g', f~YsY
Подобного рода сигналы широко
использовались для исследования
значения временного и пространственного
механизмов в восприятии высоты сложного
звука (Schouten et al., 1962; см. соответствующий
раздел главы «Психоакустика»).
В
условиях воздействия такого сложного
раздражения, в котором могут изменяться
соотношения его временных и спектраль-
299
пых
компонентов^ наблюдаются реакции
отдельных нейронов, которые отражают
сочетанное действие этих компонентов.
Формы ответа отдельных нервных единиц
на это сочетанное раздражение достаточно
разнообразны. Можно упомянуть о некоторых
характерных случаях ответа нейронов
высших отделов слуховой системы
(заднего двухолмия и слуховой коры) на
амплитудно- модулированные звуки. Так,
при использовании низких частот
модуляции (до 20—50 гц) и высоких частот
несущей, лежащих в пределах оптимальной
частоты нейрона и вне этого предела,
можно наблюдать характерные ответы на
сочетание спектральных и временных
признаков сигнала, различные для разных
нейронов. В определенных условиях могут
возникать ответы нейрона, которые
оказываются сдвинутыми по фазе на 180
град. Наблюдается также воспроизведение
временных свойств сигнала, выражаемое
в длительном послеразряде лишь при
определенных сочетаниях частоты
модуляции и частоты несущей (Гершуни,
Вартанян, 1969). Хотя факторы, определяющие
возникновение подобного рода реакций,
еще недостаточно изучены, само явление
отражает механизм выделения нейронами
центральных отделов определенных
сочетанных признаков сложного звука.
Следует
указать, что понятие сложного звука
при приложении его к слуховой системе
не ограничивается лишь формальной
характеристикой параметров звука,
но требует учета рецепторной поверхности,
на которую этот звук должен воздействовать.
Так, например, тональная посылка,
излучаемая источником звука, расположенным
в свободном звуковом поле несимметрично
относительно головы исследуемого
животного, воздействует со сдвигом
по фазе на рецепторные поверхности
правого и левого уха.
С
точки зрения слуховой системы подобный
звук является сложным звуком со
сдвинутыми по времени компонентами в
точках пространства, соответствующих
расположению правого и левого уха.
Выше
при описании частотнопороговых кривых
(рис. 72) указывалось, что установление
этих кривых осуществляется в условиях
воздействия звука на разные участки
рецептивного поля нервной единицы. Там
же обращалось внимание, что наиболее
прямым путем изучения обострения
частотной избирательности нервной
единицы, зависящим от развития торможения,
является использование звуков, состоящих
из нескольких компонентов (не менее
двух), отличающихся по спектральному
составу и воздействующих на разные
участки рецептивного поля единицы
(рис. 102). Действие такого рода сложного
звука рассматривалось как раздражение,
наиболее соответствующее свойствам
слуховой системы уже на уровне слухового
нерва. На этих предпосылках основывалась
попытка математического выражения
действия сложного звука, состоящего
из двух спектрально отличных компонентов
на рецептивном поле волокна слухового
нерва кошки (Sachs, 1969).
300
Выше
при рассмотрении вопроса о выделении
временных признаков сигнала
указывалось, что каждый сигнал обладает
временными признаками и переходными
явлениями при включении и выключении,
которые могут рассматриваться при
наличии стационарного участка
сигнала как компоненты, составляющие
звук. С этой точки зрения большинство
звуков имеет черты, позволяющие
приближать их к типу сложных звуков.
ОРГАНИЗАЦИЯ
РЕЦЕПТИВНЫХ ПОЛЕЙ ОТДЕЛЬНЫХ НЕРВНЫХ
ЕДИНИЦ
Под
рецептивным полем понимают участок
рецепторной поверхности, с которой
функционально связана нервная единица.
Установление и исследование свойств
рецептивного поля в таких сенсорных
системах, как зрительная и сомато-сенсорная,
осуществляется в условиях перемещения
раздражителя по поверхности сетчатки
или кожи. Перемещение воздействующего
раздражения по рецепторной поверхности
улитки не может наблюдаться непосредственно.
О нем можно судить лишь косвенно по
ответам нервной единицы на разные
звуковые частоты, исходя из представлений
о пространственной проекции частот на
этой поверхности.
Приведенные
выше данные о частотнопороговых и
частотноравноимпульсных кривых и
боковых тормозных зонах рассматривались
с точки зрения пространственной
организации рецептивного поля
нервной единицы слуховой системы
подобно единицам зрительной и
сомато-сенсорной систем. С этой точки
зрения тормозное взаимодействие,
наблюдаемое при двух звуковых посылках
разных частот, рассматривалось как
выражение латерального торможения,
возникающего в результате воздействия
раздражителей на центральные и
боковые участки рецептивного поля
нейрона, так же как это имеет место
в зрительной и сомато-сенсорной системах.
Вместе с тем в предыдущем изложении
было приведено большое количество
фактов, которые показывали значение
другого фактора, помимо пространственного,
а именно временных признаков и
временных характеристик возбуждающих
и тормозных явлений в деятельности
нервной единицы. Такие явления, как
развитие торможения, определяемого
интервалами между раздражениями,
наносимыми на рецепторные поверхности
обоих ушей (см. главу «Пространственный
слух»), или длительностью сигнала (рис.
106), повышение порогов при возрастании
длительности сигнала могут служить
достаточно выразительными примерами
развития тормозных явлений, которые
не сводимы к случаю пространственно
обусловленного латерального торможения.
Эти явления служат примерами торможения,
обусловленного временными факторами
структуры сигнала.
Таким
образом, в отличие от зрительной и
сомато-сенсорной систем организация
рецептивного поля единицы слуховой
системы
301
Должна
обязательно описываться сочетанием
временных и пространственных
факторов, определяющих взаимодействие
элементов рецептивного поля.
Следует
указать, что до сих пор принципы
временно-пространственной организации
рецептивного поля единицы слуховой
системы еще недостаточно изучены.
10
20 30 t 50 60 70~~ ^8096
Рис.
106. Динамика импульсной активности
нейрона заднего двухолмия
(по:
Марусева, 1969).
Графики
слева —
зависимость числа импульсов от
интенсивности звуковых посылок (белого
шума) различной длительности. Черные
кружки —
при длительности сигнала 50 мсек.,
крестики
—
5 мсек., белые
кружки —
2 мсек. По
оси абсцисс —
интенсивность в дб над уровнем 0.0002
микробара; по
оси ординат —
количество импульсов в 10 реакциях
(10п). Справа
—
кадры записи импульсных разрядов
методом точек для трех длительностей
звука (2, 5 и 50 мсек.) при разных интенсивностях
сигнала. На каждом кадре горизонтальный
ряд точек
соответствует разряду импульсов при
одном раздражении; первая
точка в ряду
показывает начало раздражения. Над
кадрами слева —
отметка длительности сигнала, цифры
над кадрами —
интенсивность сигнала в дб.
ОРГАНИЗАЦИЯ
СОВОКУПНОСТЕЙ ЕДИНИЦ
При
рассмотрении вопроса об организации
совокупностей единиц слуховой
системы следует указать, что выше уже
приводилось достаточно данных, которые
свидетельствовали, что
302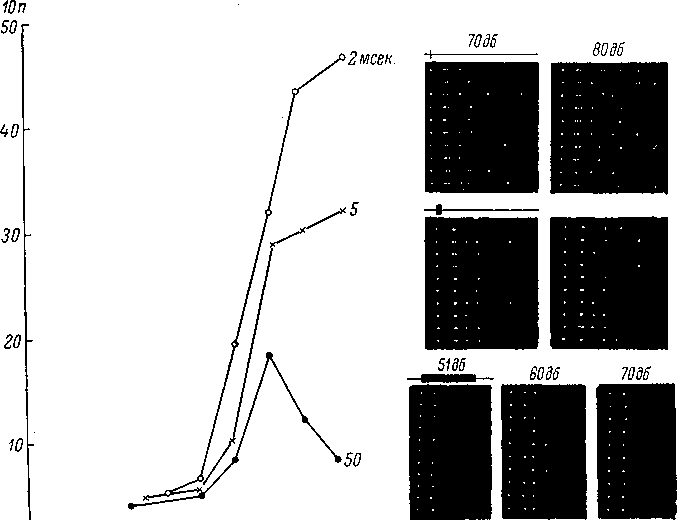
пространственная
проекция частот, начинающаяся с
поверхности улитки, многократно
представлена в центральных отделах
слуховой системы в упорядоченном
согласно частотной шкале распо-
Рис.
107. Схема проекций частот на рецепторной
поверхности улитки
и ее связей с
кохлеарными ядрами (составлена Г. И.
Ратниковой,
см.: Гершуни, 1967).
Внизу
—
поверхность развернутых завитков
улитки кошки (справа
от нее — шкала длины улитки в мм); цифры
— частоты в кгц; вверху —
кохлеарные ядра: а
— переднее вентральное, р
—
заднее вентральное, d
—
дорсальное.
ложении
нервных единиц с разными характеристичными
частотами. Однако это явление
пространственного распределения единиц
слуховой системы не только с
функциональной, но и со структурной
точки зрения еще недостаточно даже для
элементарного представления об
организации функции этой системы. Так,
уже на уровне
303
кохлеарных
ядер наличие нервных волокон, связанных
с определенными участками улитки и
расположенных друг относительно друга
после их бифуркации в дорсо-вентральном
направлении,
Рис.
108. Схема локализации точек отведения
на сагиттальном срезе
заднего холма
(по: Гершуни и др., 1969).
Кружки,
треугольники, крестики —
точки отведения по ходу электродов,
соответствующие нейронам с разным
типом реакции. Черные
кружочки —
коротколатентные, быстр ©суммирующие
фазические нейроны; черные
треугольники — длиннолатентные,
медленносуммирующие тонические нейроны;
полые
кружки и
треугольники,
а
также крестики
—
промежуточные группы. Цифры
обозначают оптимальные частоты,
соответствующие точкам отведения и
номера проколов (1—7).
На рисунке видны нейроны с резко
отличными временными характеристиками
как по ходу частотной шкалы (проколы
4,
5),
так и для изочастотных групп (проколы
I, 7).
определяет
частотную проекцию, а ход отдельного
волокна в сагиттальной плоскости дает
развертку каждого участка улитки во
всех отделах кохлеарного ядра (рис.
107). Таким образом, одновременно с
частотной проекцией в объеме слуховой
системы
304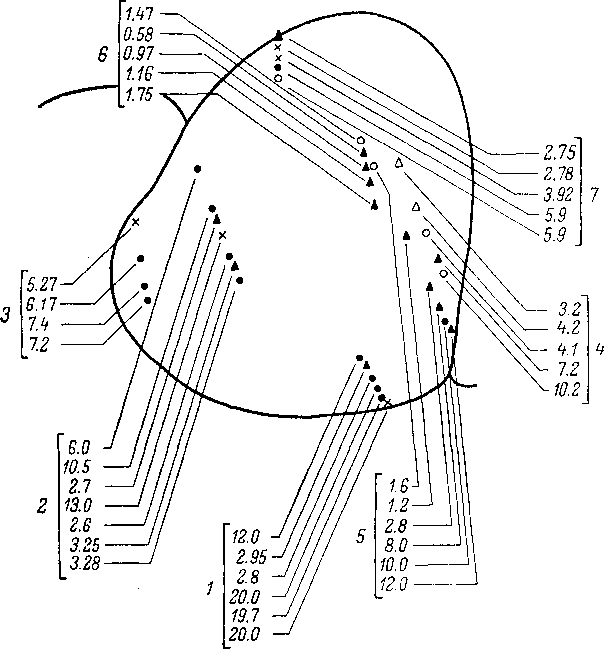
должна
осуществляться упорядоченная проекция
одних и тех же участков улитки, которую
можно обозначить как изочастот- ную.
Исследование
свойств нейронов, обладающих одной и
той же или близкой характеристичной
частотой как в кохлеарном ядре, так и
в заднем двухолмии, показывает, что в
составе изочастот- ной проекции (рис.
108) имеются нейроны, обладающие резко
отличными временными характеристиками.
Таким
образом, приходится признать, что
сведения, поступающие от одного и
того же участка улитки, должны
обрабатываться по шкале времени
по-разному, различными группами
изочастотных элементов. В целом очевидно,
что в центральных отделах описание
свойств сигнала осушествляется
одновременно действующими группами
элементов, обладающими отличными
возможностями передачи сведений о
различных признаках (компонентах)
сигнала.
Все
изложенное выше позволяет в самом общем
виде охарактеризовать слуховую
систему как систему, во-первых, выделяющую
сведения об отдельных параметрах и
компонентах звукового сигнала и,
во-вторых, воспринимающую специфическим
образом определенные^сочетания этих
параметров и компонентов.
Как
излагалось ранее (см. главу «Общие
характеристики слуха позвоночных»),
естественными звуками, к восприятию
которых в процессе эволюции позвоночных
животных приспосабливалась слуховая
система, являлись сложные звуки
биологического происхождения. Там
же приводились примеры исследования
реакций определенных нервных элементов
слухового нерва на сложные звуки у
амфибий (лягушки), свидетельствующие
о наличии явлений тормозного взаимодействия
сходного с подобными же явлениями,
наблюдаемыми у млекопитающих при
действии двух отличных по своим
спектральным признакам компонентов
сложного звука (Frishkopf, Goldstein, 1963; Бибиков,
1969, 1970).
Очевидно,
что свойство выделять различные
параметры звука и реагировать на
определенные сочетания этих параметров
и таким образом воспринимать звуки,
состоящие из разных компонентов (т. е.
сложные звуки), является общим свойством,
характерным для слуховой системы
позвоночных.
Рассмотрение
с изложенной точки зрения восприятия
любого звукового сигнала, начиная с
одиночных звуковых посылок определенной
частоты и минимальной интенсивности,
используемых при пороговых измерениях,
показывает, что и в этом случае, в
определенных отношениях наиболее
простом, в слуховой системе происходит
передача сведений о разных признаках
сигнала (уровне интенсивности,
спектральной характеристике, временных
свойствах), т. е. так же, как это имеет
место при восприятии сложных звуков.
20
Сенсорные системы
305
На
основании всего изложенного становится
очевидным, что реально не существует
сигналов, которые обладали бы лишь
одним параметром, но существуют
физические приборы, которые измеряют
один параметр, например частоту
стационарного периодического
колебания синусоидальной формы. Слуховая
система не может быть аналогизирована
с таким прибором, ибо она всегда
осуществляет измерение целого ряда
параметров звука.
Следует
подчеркнуть также, что слуховая система
при любом способе ее исследования,
включая изучение свойств отдельных ее
единиц и в любых условиях раздражения,
не может рассматриваться с точки
зрения функции изолированных, не
взаимодействующих друг с другом
элементов, как это постулировалось
классической теорией прошлого века.5
Однако степень и характер этого
взаимодействия, в первую очередь
тормозного, может быть иным в случаях
действия разных сигналов, с одной
стороны, и разных задачах, осуществляемых
слуховой системой при их обнаружении,
различении и опознавания — с другой.
Поэтому
каждый случай восприятия звуков, который
раздельно рассматривается психоакустикой
(например, измерение абсолютных
порогов, дифференциальных порогов,
маскировка, восприятие сложных
звуков) должен характеризоваться
особенностями действия
нейрофизиологических механизмов, и
хотя эти особенности до настоящего
времени еще совершенно недостаточно
изучены, они все же требуют упоминания.
Так,
особенностью случая пороговых измерений
интенсивности является осуществление
деятельности в условиях раздражения
минимального числа элементов рецепторной
поверхности и связанных с ними волокон
слухового нерва. Уже в этих условиях
слуховой системой резко отделяется
начальный момент действия звука от
последующего и выделяются спектральные
свойства сигнала, как показывает снятие
частотнопороговых кривых по первому
импульсу коротколатентных единиц в
кохлеарных ядрах и особенно в заднем
двухолмии.
Наиболее
быстрая передача этих сведений о
свойствах сигнала, содержащаяся в
начальных импульсах волокон слухового
нерва, обеспечивается функциональной
организацией всех вышележащих
отделов слуховой системы. Эта
функциональная организация, выступающая
наиболее отчетливо в условиях восприятия
коротких околопороговых звуков,
характеризуется наименьшим взаимодействием
нервных единиц в нижележащих отделах
слуховой системы, что, в частности,
выражается в отсутствии тормозных
влияний для очень коротких звуков на
уровне задних холмов (рис. 106) и резким
его возрастанием на корковом уровне.
Эти данные дают основание полагать,
что при действии
5
См. Предисловие к первой части тома
«Физиология сенсорных систем».
<<Наука», Л., 1971.
30§
коротких
слабых звуков основная конвергенция
импульсой должна осуществляться на
нейронах коркового уровня.
Очевидно,
что подобное малое взаимодействие
единиц на нижележащих уровнях и
обеспечение его на корковом уровне при
действии коротких звуков может являться
одним из факторов, определяющих нарушение
восприятия коротких звуков именно при
повреждении слуховой коры, а не нижележщих
отделов слуховой системы (о значении
механизма краткострочной памяти см.
главу «Роль различных отделов слуховой
системы в восприятии звуков»).
В
целом имеющиеся в настоящее время
данные о нейрофизиологических
механизмах операций, осуществляемых
слуховой системой при восприятии
звуков, еще недостаточны для построения
общей нейрофизиологической теории.
Поэтому в настоящем руководстве данные
психофизического исследования слуха
не могли быть систематически
рассмотрены с точки зрения лежащих в
их основе нейрофизиологических
механизмов. Можно полагать, что подобное
рассмотрение благодаря быстрому росту
знаний станет возможным в недалеком
будущем.
.20*