

ВЕСТИБУЛЯРНАЯ
СИСТЕМА
Вестибулярный
аппарат является многомерным
биологическим преобразователем
механической энергии угловых и линейных
ускорений в сигналы о положении и
движении тела. Его многомерность
обеспечивается наличием расположенных
в разных плоскостях трехмерного
пространства трех пар полукружных
каналов, пары утрикулюсов и пары
саккулюсов. Вестибулярный аппарат
связан с нервными центрами и эффекторными
органами, образуя сложную систему
управления положением головы и тела в
пространстве. Эту функцию вестибулярная
система выполняет во взаимодействии
с другими сенсорными системами
организма.
Вестибулярный
аппарат — часть внутреннего уха
(лабиринта), расположенного в толще
каменистой части височной кости, состоит
из костного и перепончатого лабиринтов.
Костный лабиринт — это система трех
туннелей — полукружных каналов (canales
semicirculares) и сообщающаяся с ними полость
— преддверие (vestibulum). Перепончатый
лабиринт — система тонкостенных
соединительнотканных трубок и мешочков,
отчасти повторяющая форму костного
лабиринта и расположенная внутри его.
В
филогенезе вестибулярный аппарат
развивается на основе органов баковой
линии. Сначала из канала органа боковой
линии образуется пузырек — общий мешок,
затем на нем формируется один полукружный
канал (у миксин), далее два (у миног) и
три (у поперечноротых). Дифференцировка
мешочка на sacculus и utriculus отсутствует у
миксины, но намечена у миноги, у которой
имеется общий мешок (saccus communis), делящийся
на две части, и два полукружных канала
— передний и задний — в виде начальных
ампулообразных расширений. Таким
образом вертикальные каналы
филогенетически являются более древними,
57Глава 3
Краткие анатомические сведения
чем
горизонтальные (Сепп, 1959). На последующих
этапах филогенеза строение
вестибулярного аппарата меняется в
деталях, но принципиально остается тем
же. В анатомии лабиринта имеются как
видовые, так и индивидуальные различия.
Сравнительноанатомическим особенностям
лабиринта посвящено значительное число
работ (Штейн, 1890; Калина, 1960; Werner, 1960, и
др.).
Плоскости
полукружных каналов расположены
примерно под прямым углом друг к другу;
полукружные каналы, лежащие в одной
плоскости или в параллельных плоскостях,
составляют функциональные пары.
Расположение каналов по отношению к
трем основным плоскостям — горизонтальной,
фронтальной и сагиттальной — хотя и
подвержено значительным вариациям,
особенно видовым, в общем сводится к
следующему. Составляющие функциональную
пару правый и левый наружные (синонимы:
горизонтальные, латеральные) каналы
лежат в плоскости, образующей с
горизонтальной плоскостью угол, примерно
равный 30 град., открытый вперед и вверх.
Плоскость переднего (верхнего или
фронтального) канала составляет с
фронтальной плоскостью головы угол
45 град., открытый латерально и вперед.
Примерно такой же величины угол, открытый
латерально и назад, имеется между
плоскостью заднего (нижнего или
сагиттального) канала и фронтальной
плоскостью. Функциональные пары
вертикальных каналов образованы
расположенными в параллельных
плоскостях передним полукружным каналом
одной стороны и задним полукружным
каналом противоположной стороны.
Наличие трех функциональных пар каналов
обеспечивает их участие в любом повороте
головы в любой плоскости. Полукружные
каналы сообщаются с полостью преддверия
посредством пяти отверстий. Тремя
из них сообщаются с преддверием
ампулярные расширения каналов.
Простые ножки (crus simplex) каналов также
сообщаются с преддверием, но через два
отверстия: через одно — наружный канал,
через другое — оба вертикальных
канала, так как простые ножки вертикальных
каналов до впадения их в преддверие
объединяются в общую ножку (crus communis).
При нормальном положении головы (теменем
вверх) передний полукружный канал
обращен дугой вверх, ампулой наружу,
а простой ножкой внутрь; дуга заднего
канала обращена назад и наружу
(ампула — вниз, а простая ножка кверху);
наружный канал обращен дугой наружу,
ампулой вперед, а простой ножкой кзади.
Внутри
костной капсулы лабиринта расположен
перепончатый лабиринт, подвешенный
к костным стенкам фиброзными перемычками
(рис. 24). В костных ампулах перепончатые
каналы расширяются. В преддверии
перепончатый лабиринт образует два
сообщающихся между собой мешочка:
утрикулюс, с которым сообщаются
перепончатые полукружные каналы, и
саккулюс. Мешочки преддверия прилежат
к медиальной стенке
58
костного
преддверия и расположены в сферическом
(саккулюс) и эллиптическом (утрикулюс)
углублениях этой стенки.
Некоторые
основные размеры (в мм) перепончатого
лабиринта
человека
приведены в табл. 2 (по: Anson et al., 1967).
Данные
о размерах лабиринта различных
позвоночных животных приведены в
ряде работ (Camis, 1930; Jones, Spells, 1963;
Рис.
24. Перепончатый лабиринт человека —
правый лабиринт, вид изнутри и несколько
сверху (по: Anson et al., 1967).
Утрикулюс
(I), саккулюс (2) и ампулы (3,
4,
5) вскрыты, чтобы показать расположение
утрикулярной (6) и саккулярной (7) макул,
ампулярных крист (8,
9, 10)
и полукружных каналов — переднего
(11),
наружного (12)
и заднего (13).
Эндолимфатический мешок (14)
показан частично. VIII чер.епно-мозговой
нерв состоит из кохлеарной (15)
и
вестибулярной (16)
частей. Последняя в свою очередь дает
ветви к ампулам верхнего и наружного
каналов (17),
к утрикулюсу (18),
к саккулюсу (19)
и к ампуле заднего канала (20).
Остальные обозначения: 21
—
лицевой нерв, 22
— спиральный ганглий кохлеарного
нерва, 23
—
кохлеарный проток, 24
— соединительный проток, 25
— эндолимфатический
проток, 26
—
общая ножка верхнего и заднего полукружных
каналов. Вверху показано схематическое
положение каналов в черепе.
Fernandez,
Valentinuzzi, 1968). Перепончатые полукружные
каналы и мешочки преддверия заполнены
эндолимфой 1
и сообщаются с улиткой, а также с
расположенным в полости черепа
эндолимфатическим мешком. Существует
мнение, что последний участвует в обмене
эндолимфы, секретируя ее (Seymour, 1954).
Эндолимфатическое пространство
анатомически замкнуто.
1 Физико-химическим
свойствам эндолимфы посвящены специальные
исследования (Maggio, 1966),
59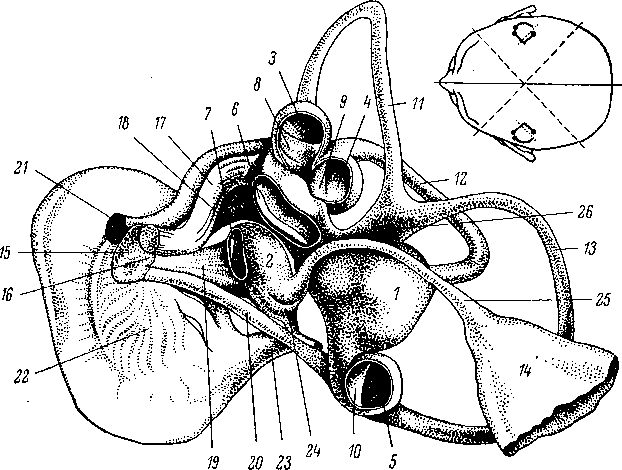
Таблица
2
Основные
размеры (в мм) некоторых частей
перепончатого
лабиринта человека |
Длина |
Ширина |
Окружность |
Утрикулюс |
4.2 |
2 |
6.5 |
Макула утрикулюса |
2.3 |
2.1 |
— |
Саккулюс |
2.6 |
2.2 |
5.4 |
Макула саккулюса ....... |
2.2 |
1.2 |
— |
Ампула полукружного канала |
1.8 |
1.5 |
4.9 |
Перепончатый
лабиринт имеет тонкую фиброзную стенку,
выстлан изнутри уплощенным эпителием
и расположен эксцентрично в костной
капсуле лабиринта. Просвет его^
заполненный эндолимфой, почти на всем
протяжении (за исключением ампул)
составляет лишь около 1/4 просвета
костных каналов, диаметр которых у
человека примерно равен 1 мм.
Перилимфатическое пространство между
перепончатым и костным лабиринтами
сообщается с подпаутинным пространством
головного мозга.
Перепончатый
лабиринт плотно прилегает к костному
только в местах концевых нервных
аппаратов, т. е. на участках костных
гребешков ампул (крист) и пятен преддверия
(макул). Плоскость утрикулярной
макулы у человека совпадает с плоскостью
наружного канала, а плоскость саккулярной
макулы перпендикулярна ей. Анатомическое
и, возможно, функциональное различие
отолитовых органов у животных разных
видов более значительно, чем различие
в полукружных каналах.
Вестибулярный
нерв образован отростками биполярных
нейронов, составляющих в совокупности
ганглий Скарна. Периферические
отростки этих клеток объединяются в
несколько ветвей, идущих к каждому
из трех полукружных каналов, к утри-
кулюсу и саккулюсу. Центральные отростки
направляются к вестибулярным ядрам
продолговатого мозга [(Brodal, 1967а).
По
данным Лоренте де Но (Lorente de No, 1931), у
мышей утрикулярные и ампулярные волокна
вестибулярного нерва связаны с
верхней половиной ганглия, а саккулярные
— с нижней. Волокна из заднего полукружного
канала идут несколько обособленно
и лишь в самом ганглии присоединяются
к волокнам из остальных полукружных
каналов. У обезьян ганглий Скарпа
состоит из верхнего вестибулярного
ганглия, снабжающего нервными
волокнами кристы наружного и переднего
каналов, а также утрикулярную макулу,
и нижнего вестибулярного ганглия,
иннервирующего кристу заднего канала
и саккулярную макулу. Зоны ганглия
Скарпа, иннервирующие кристы полукружных
каналов, снабжают волокнами главным
образом верхнее вестибулярное
60
ядро
и оральную часть медиального вестибулярного
ядра. Зоны, иннервирующие утрикулярную
макулу, проецируются на^ме- диальное и
нижнее вестибулярные ядра, а центральные
волокна саккулярной части ганглия идут
к дорсо-латеральной области нижнего
вестибулярного ядра. Проекций к
латеральному ядру у обезьян не обнаружено,
и полагают, что вестибулярные импульсы
поступают к нему через мозжечок (Stein,
Carpenter, 1967). Однако у кошек первичные
вестибулярные волокна снабжают
ростровентральную область латерального
ядра (Brodal et al., 1962).
Волокна
вестибулярного нерва составляют
значительную часть VIII черепномозгового
нерва (n. stato-acusticus). У человека площадь,
которую занимают чувствительные клетки
вестибулярного аппарата, составляет
11.27 мм2,
количество миелиновых волокон в
вестибулярном нерве в среднем 19 000,
диаметр волокон — от 1 до 13 мкм
(Блинков, Глезер, 1964).
ФИЗИОЛОГИЯ
РЕЦЕПТОРОВ ЛАБИРИНТА
Вестибулярный
сенсорный эпителий. Современное
представление об организации
вестибулярного сенсорного эпителия
сложилось прежде всего благодаря
фундаментальным исследованиям Яна
Вёрселла и его сотрудников (Wersall, 1956;
Engstrom, 1958; Engstrom, Wersall, 1958; Flock, Duvall, 1965, и
др.). Вестибулярные сенсорные клетки
млекопитающих образуют 5 эпителиальных
областей: по одной в каждой ампуле
полукружных каналов (crista ampullaris) и по
одной — в утрикулюсе и саккулюсе (macula
utriculi и macula sacculi). Сенсорные клетки
дифференцируются на два типа (рис. 25),
причем у животных, стоящих на более
низких уровнях эволюционного ряда
(рыбы, амфибии), можно обнаружить лишь
относительно более простые клетки
второго типа (Wersall, 1956).
Клетки
типа I имеют форму колбы с закругленным
дном и размещены в бокалообразной
нервной полости. Обычно каждая полость
содержит лишь одну сенсорную клетку,
однако в утрикуляр- ной и сакцулярной
макулах можно обнаружить в одной полости
2—4 сенсорные клетки (Engstrom, 1961). В
утрикулярной макуле птиц это число
может составлять 5—7 (Винников, 1966).
Нижняя часть сенсорной клетки отделена
от нервной капсулы межклеточным
пространством правильной формы, толщиной
150—200 А. Область горловины клетки
отделена от нервной капсулы более узким
(100 А) межклеточным пространством
(Engstrom, Wersall, 1958).
Клетки
типа II имеют вид цилиндров с округлым
основанием. В разных частях crista ampullaris
встречаются как длинные тонкие,, так
и короткие толстые клетки типа II.
Характерная особенность этих клеток
заключается в _их иннервации: нервные
61
окончания
здесь могут быть как афферентными
(большинство), так и эфферентными
(Engstrom, Wersall, 1958). Кроме клеток двух основных
типов, существуют клетки промежуточного
типа. У животных, в вестибулярном
эпителии которых встречаются оба типа
клеток, клетки типа I обычно локализуются
в центре crista ampullaris, тогда как клетки
типа II — на ее периферии (Титова,
1968).
Рис.
25. Схематическое изображение вестибулярных
сенсорных клеток
типа I (НС
I)
и II (НС
II),
показывающее сложность их нервных
связей
(Ades, Engstrom, 1965).
Клетки
обоих типов могут контактировать с
одной и той же нервной полостью (NC).
Боковая
веточка из нервной полости клетки типа
I может контактировать с клеткой типа
II. Показаны различные формы нервных
окончаний двух типов (NEi
и NE2).
НС —
киноцилии, Н
—
стереоцилии.
Первоначальное
предположение о том, что два типа
сенсорных клеток могут иннервироваться
волокнами разных типов (Wersall, 1956; Engstrom,
Wersall, 1958), оказалось в определенной мере
правильным, однако позднее выяснилось,
что иннервация сенсорных клеток в
действительности выглядит гораздо
сложнее. В частности, имеет место
перекрытие иннервации, так что клетки
типов I и II могут иннервироваться одним
волокном. Внешняя
62
поверхность
нервной полости клеток типа I часто
образует контакт с клетками типа II
(рис. 25) — либо непосредственно, либо с
помощью коллатералей (Ades, Engstrom, 1965).
Клетки
обоих типов несут пучок волосков,
выступающих из кутикулы на их верхушках.
Каждый пучок насчитывает 50— 80 волосков
(цилий) в клетках крист (Wersalll, 1956) и 40—110
ци- лий — в клетках макул (Spoendlin, 1964). В
каждом пучке, кроме стереоцилий, имеется
одна киноцилия 2
— более длинный отросток, расположенный
всегда на периферии пучка. Расположение
стереоцилий таково, что самые длинные
из них находятся вблизи киноцилии,
а длина остальных прогрессивно
уменьшается по мере удаления от
киноцилии. Такой «лестничный» характер
расположения стереоцилий относительно
киноцилии является типичным.
В
последнее время в сенсорном эпителии
отолитовых органов лягушки между
киноцилией и пятью прилежащими
стереоцилиями обнаружен соединительный
комплекс в виде нитеподобных структур.
Эти нити пронизывают межцилйарное
пространство (около 1200 А) и, достигнув
мембраны стереоцилий, веерообразно
расходятся. Морфологическая связь
между кино- и стереоцилиями указывает
на существование структурной единицы
— цилиарного аппарата, который может
быть первым звеном в цепи преобразования
информации: сенсорный эпителий — центры
(Hillman, 1969).
Вестибулярный
эпителий всех пяти сенсорных областей
имеет определенную морфологическую
поляризацию. Например, в кристе
горизонтального канала киноцилии
волосковых клеток обращены к утрикулюсу,
а в кристе переднего вертикального
канала они всегда обращены к просвету
канала. По всей вероят- сти, это имеет
отношение к механизму дирекционной
чувствительности каналов, поскольку
и в горизонтальном, и в вертикальных
каналах возбуждающий ток эндолимфы
направлен от стереоцилий к киноцилиям,
а тормозящий — наоборот (Lowenstein,
Wersall, 1959). Несколько иначе расположены
киноцилии в эпителии макул: здесь
сенсорные клетки образуют группы из
нескольких сот единиц. Внутри каждой
такой группы киноцилии ориентированы
одинаково, однако ориентация самих
групп различна (Flock, 1964; Spoendlin, 1964). При
реконструкции поверхности макул с
помощью горизонтальных сериальных
срезов было обнаружено (Spoendlin, 1965), что
линии поляризации веерообразно
расходятся от медиальной передней
части макулы к изогнутой границе,
за пределами которой поляризация
реверсирует (рис. 26).
2 Следует
отметить, что у беспозвоночных (в
частности, осьминогов и кальмаров) на
каждую рецепторную клетку приходится
не одна, как у позвоночных, а 70—120
киноцилий (Винников и др., 1968).
63
Волоски
эпителиальных клеток кристы покрыты
желеобразным колпачком (cupula
terminalis). Волоски чувствительных клеток
макул погружены в желеобразную мембрану,
содержащую кристаллические включения
отолиты. С помощью методов электронной
микроскопии и дифракции рентгеновских
лучей установлено, что эти кристаллы
у человека и у морской свинки состоят
из кальцита плотностью 2.71 (Carlstrom et al.,
1953).
По
поводу механизма функционирования
волосковых клеток вестибулярного
эпителия в настоящее время выдвинуты
две гипотезы, ни одну из которых
нельзя считать доказанной. Согласно
одной из них, волосковая клетка действует
по принципу конден-
Рис.
26. Схематическое изображение
поляризации
сенсорных клеток в
macula utriculi морской свинки
(Spoendlin, 1965).
Стрелки
указывают направление поляризации,
веерообразно расходящейся от медиального
края макулы до линии (пунктир),
за которой знак поляризации изменяется.
Штрихпунктирными
линиями
показаны сагиттальная и фронтальная
оси.
сатора:
положительный заряд, приложенный с
внешней стороны клеточной мембраны,
вызывает электронегативность растворов
клеточной протоплазмы. При этом
допускается, что клеточная мембрана
непроницаема для ионов калия (Dohlman,
1960а), содержание которого в эндолимфе
велико и составляет около 140 мэкв/л
(Smith et al., 1958), в противоположность низкому
содержанию натрия (Citron et al., 1956). Согласно
второй точке зрения, мембрана проницаема
для ионов калия, и проницаемость (а
значит и чувствительность волосковой
клетки) управляется потенциалами
смещения гиалуронатов (Christiansen, 1963).
Гиалуронаты — соли гиалуроновой
кислоты, относящиеся к мукополисахаридам,
вещества с высоким (около 106)
молекулярным весом — были обнаружены
в лабиринте гистохимическими методами
(Vilstrup, Jensen, 1954). Калиевые соли гиалуроновой
кислоты образуют с водой вязкие
растворы. Работами Дольмана (Dohlman, 1959 и
др.) было показано, что в crista ampullaris 64
вокруг
сенсорного эпителия имеется особая
область — planum semilunatum, секретирующая
сульфомукополисахариды. При деформации
молекул этих веществ возникает
электрохимический «потенциал смещения»,
который, по-видимому, имеет прямое
отношение к функции волосковой клетки.
Это название связано
с
характером эксперимента, в котором
раствор гиалуроната калия,
смещаясь
в капилляре, создавал разность потенциалов
(до 110 мв)
между
двумя опущенными в раствор электродами.
Изменения сме-
щения столбика жидкости
отражались в соответствующих изме-
нениях
регистрируемого потенциала,
дить
широкий диапазон звуковых
частот
(Vilstrup, Jensen, 1961).
Наличие
в лабиринте (как в слу-
ховой, так и
в вестибулярной его
частях) веществ
медиаторного типа,
прежде всего
ацетилхолина (см. об-
зор: Trincker, 1959),
приводит к пред-
положению о
существовании меха-
низма усиления
вестибулярных им-
пульсов (Lowenstein,
1961).
Измерения
постоянных потенциа-
способного
воспроизво-
Рис.
27. Распределение по-
стоянных
потенциалов в ку-
пуло-эндолимфатическом
про-
странстве и кристе полукруж-
ного
канала морской свинки
(по: Trincker,
1957).
Измерения
производились относительно перилимфы.
Плюсы
и минусы
соответствуют знаку потенциалов.
Остальные объяснения в тексте.
лов
в crista ampullaris, купуле, пе-
ри- и
эндолимфатическом пространст-
ве в
покое и при адекватной стиму-
ляции
(Trincker, 1957) заложили
основу представлений
о механизме
вестибулярного возбуждения.
Рас-
пределение постоянных
потенциалов
дано на рис. 27. Оказалось,
что
величины
потенциалов покоя полу-
кружного
канала в общем соответст-
вуют
измерениям Бекеши (Bekesy, 1952), проведенным
на улитке.
Потенциалы кристы и купулы
также имеют характерное распре-
деление:
максимальный потенциал поверхности
купулы достигает
почти +80 мв, кристы
— почти —60 мв (относительно перилимфы).
При
отклонениях купулы от положения покоя
в сторону утрику-
люса
в горизонтальном канале возникала
деполяризация, а в переднем вертикальном
канале — гиперполяризация. При утрику-
лофугальных отклонениях купул происходили
обратные изменения. Диапазон
физиологически важных отклонений
купулы (в пределах 30 град, около нулевого
положения) соответствует линейной
части S-образной кривой, выражающей
соотношение угла отклонения купулы и
сдвига постоянных потенци
алов.
Работы
Тринкера (Trincker, 1957) и более ранние
исследования Лёвенштейна (Lowenstein, 1955)
позволяют говорить о существовании
в волосковых клетках рецепторного
потенциала — свя-
5
Сенсорные
системы
65
дующего
звена между действием механического
стимулй й изменениями частотного
кода в вестибулярном нерве. Согласно
современным представлениям,
рецепторный потенциал, вызванный
смещением купулы относительно кристы,
в свою очередь модулирует частоту
импульсов в вестибулярном нерве путем
де- или гиперполяризации дендритных
нервных окончаний в области синапсов
с телами волосковых клеток (Lowenstein,
1961).
Причины
существования постоянных потенциалов
в лабиринте достоверно не известны. По
мнению Бекеши (Bekesy, 1951), это связано с
разностью концентраций ионов, в
особенности К+.
Однако позднее было обнаружено, что
хотя постоянные потенциалы улитки
примерно на 70 мв выше, чем потенциалы
утрикулюса и саккулюса, высокое
содержание калия и низкая концентрация
натрия в эндолимфе всюду одинаковы. На
этом основании делается вывод о том,
что эндолимфатические потенциалы покоя
внутреннего уха не связаны с содержанием
калия и натрия в эндолимфе (Smith et al.,
1958).
Функция
полукружных каналов. Теория купулы.
Способы стимуляции и пороги чувствительности
полукружных каналов. Современная
характеристика функций полукружных
каналов стала возможной после работ
Штейнгаузена (Steinhausen, 1931, 1933), разработавшего
метод окраски и прижизненного наблюдения
купулы в полукружном канале щуки.
Позднее * это удалось повторить и
другим исследователям (Dohlman, 1935; Vilstrup,
1950; Trincker, 1957). Купула в оптическом
отношении не отличается от эндолимфы;
форму и положение купулы обнаруживали
благодаря нанесению на ее поверхность
очень мелких частиц туши. Техника
Штейнгаузена позволила осуществить
фото- и кинорегистрацию отклонений
купулы и доказать, что адекватным
стимулом для купулы является угловое
ускорение и связанные с ним силы
инерции. Регистрация отклонений купулы
дала возможность сопоставить их с
движениями глаз. Тогда же была обоснована
точка зрения, согласно которой система
купула—эндолимфа] представляет собой
торсионный маятник с высокой степенью
затухания и большой инерцией. После
внезапной остановки вращения
возникает инерционный сдвиг эндолимфы,
под влиянием которого происходит
изгибание купулы, почти полностью
перекрывающей просвет ампулы. Достигнув
максимального отклонения, купула при
отсутствии дальнейших внешних воздействий
будет стремиться вернуться в исходное
(нулевое) положение в силу своих
эластических свойств. Аналогичный
механизм имеет место при любых стимулах,
результатом которых является возникновение
разности давлений эндолимфы на
противоположные поверхности купулы
(положительные и отрицательные ускорения,
калорическая проба и др.). Время
возвращения купулы к нулевой точке
колеблется в пределах от 1 до 60 сек., в
зависимости от величины предшествующего
ускоре
66

ния.
Длительность постощущения вращения
таким образом может быть связана со
временем возвращения купулы.3
Наблюдения
Штейнгаузена составили основу
купулометрии — направления, развитого
затем Утрехтской физиологической
школой (Egmond et al., 1949, 1952; Egmond, Groen, 1955; Groen
et al., 1952; Groen, 1957, 1961a, и др.). В купулометрических
исследованиях используются более
слабые стимулы, чем при вращательной
пробе Барани. Постоянная скорость (1—60
град./сек.) набирается с подпороговым
ускорением. Затем, после некоторого
периода вращения с постоянной скоростью,
производится внезапное торможение
(стоп-момент, длящийся 1—3 сек.). При этом
продолжительность постнистагма или
иллюзии противовращения является
функцией достигнутой угловой скорости.
Эта зависимость, выраженная графически,
носит название купулограммы. Данные
измерений наносятся на полулогарифмическую
шкалу: по оси абсцисс откладывают
угловые скорости кресла перед началом
торможения, по оси ординат — длительность
постнистагма или иллюзии противовращения.
В норме данные укладываются на некоторую
прямую, наклон которой не изменяется
при колебаниях порога возбудимости и
обусловлен чисто механическими
свойствами движения эндолимфы.4
Согласно Гроену и сотр., сравнение
графиков продолжительности ощущения
и нистагма давало гораздо лучшие
показания отклонений вестибулярной
функции от нормы, чем ранее применявшиеся
методики.
С
помощью резонансных колебаний на
торсионных качелях (в горизонтальной
плоскости) была определена собственная
частота купуло-эндолимфатической
системы. Если частота колебаний
качелей слишком велика, то ощущение
покоя в точке поворота качелей
отстает; когда же частота слишком мала,
ощущение опережает движение качелей.
Для случая резонансных колебаний
3 Попытки
непосредственного определения удельного
веса купулы полукружного канала до
сих пор успехом не увенчались. Однако
некоторые величины, характеризующие
физические свойства купулы, в том числе
и удельный вес, были получены в
экспериментах на боковой линии ершей,
где удельный вес купулы оказался
равным 1.01 г/см3
(возможная ошибка метода 0.002), а жесткость
— 2.5—10.0 дин/см (Jielof et al., 1952). Ввиду
большого сходства строения полукружных
каналов и органов боковой линии эти
данные могут быть отнесены и к купулам
полукружных каналов.
4 В
последнее время были определены удельный
вес и вязкость эндолимфы и перилимфы
голубя. При 40° С (температура тела
голубя) удельный вес эндолимфы равен
1.0033, а перилимфы 1.0022 г/см3
(за единицу принят удельный вес воды
при 4°). Вязкость эндолимфы равна 1.15
спуаз, а вязкость перилимфы — 0.78 спуаз
(Money et al., 1966).
Имеются
также сведения о физических константах
эндолимфы и перилимфы человека.
Константы эндолимфы: удельный вес 1.00
г/см3,
коэффициент расширения 4.4Х 10~4/°С,
вязкость 0.852 спуаз, удельная вязкость
1.18, температурный коэффициент вязкости
—2.4%/°C. Для перилимфы соответственно:
1.00 г/см3;
4.4Х10’4/°С;
0.802 спуаз; 1.11;* — 2.3%/°С (Steer et al., 1968). В тойг
же работе приведены данные о физических
константах перилимфы кошки.
5*
67
ощущение
соответствует движению качелей.
Найденная Гроеном и сотр. величина
собственной частоты дала возможность
определить константы дифференциального
уравнения маятника для купуло-эндолимфатической
системы.'5
Оценивая
значение купулометрии и отмечая ее
строгое физическое обоснование,
следует сказать о важной роли, которую
ку- пулометрия сыграла в изучении
функции полукружных каналов. В
практическом отношении купулометрия
во многих случаях является адекватным
методом в том смысле, что она позволяет
оценивать чувствительность к
укачиванию и другим видам вестибулярных
раздражений, результатом которых может
быть болезнь движения. Однако недавно
высказано мнение о том, что купулометрия
в гораздо большей степени приемлема
для анализа активности изолированного
лабиринта (Groen et al., 1952), чем для целого
организма, поскольку этот метод в его
сегодняшнем виде не учитывает эфферентной
регуляции чувствительности вестибулярных
рецепторов (Groen, 1961а), а также не всегда
позволяет получить удовлетворительные
результаты без предварительной
тренировки испытуемых.
Таким
образом, можно считать твердо
установленным, что в основе функционирования
полукружного канала лежит отклонение
купулы от положения равновесия под
влиянием любого ускорения достаточной
величины, имеющего составляющие в
плоскости данного канала.
Для
создания широкого диапазона ускорений
и скоростей в последнее время
сконструированы различные виды
вращающихся стендов с электронным
управлением (Миньковский, 1959, 1968;
Маркарян, Щукин, 1966; Черноруцкий, Сибрин,
1966). Некоторые из этих стендов
допускают возможность получения
синусоидальных колебаний скорости
по заданному закону, подобных тем,
которые ранее создавались с помощью
торсионных качелей (Egm on d et al., 1949; Groen et
al., 1952, и др.). Применение вращательных
тестов в сочетании с методом
электронистагмографии позволило
определить пороги чувствительности
полукружных каналов для угловых
ускорений. Разные авторы приводят
различные значения этих порогов. В
работах, проведенных на голубях
(Buys, Rylant, 1939; Eyck et al., 1957), пороговые
ускорения, зарегистрированные по
появлению шейного нистагма, равны
соответственно 0.8 и 1.5 град./сек.2
Для кроликов эта величина составляет
0.1—1.5 град./сек.2
(Groen, Jongkees, 1948). Тоническое отклонение
глазу людей возникает при ускорении
0.2— 0.3 град./сек.2
(Cogan, 1956), а для появления нистагма необхо
5 Это
уравнение для полукружного канала
ската имеет вид: Ё
+35$ (Groen et al., 1952). Позднее были получены
уравнения полу
кружных
каналов для лягушки (Hartog, 1963), голубя
(Money et al., 1966) и щуки (ten Kate, 1969).
68
димы,
по-видимому, большие величины ускорений:
1 град./сек.2
(Mittermaier,
Rossberg, 1956) или даже 6.4 град./сек.2
(Миньков- ский, 1966).
Что
касается величин пороговых угловых
ускорений, вызывающих субъективные
реакции (ощущение вращения, окуло-
жиральная иллюзия), то имеющиеся данные
характеризуются пестротой, отчасти
объяснимой разнообразием условий
эксперимента и применявшихся методик.
Во всяком случае в подробной сводке по
этому вопросу (Clark, 1967) приведены величины
от 0..035 до 8.2 град./сек.2
Специальное исследование вопроса о
пороге восприятия вращения, проведенное
с соблюдением всех требований (высокая
точность измерений, включая контроль
ускорений с точностью до 0.01 град./сек.2,
случайная последовательность
стимулов, большое количество испытуемых),
дало гораздо меньший разброс — от 0.05
до 2.20, в среднем 0.41 град./сек.2
Для
окуложиральной иллюзии соответствующие
величины составляют 0.04 и 0.28, в среднем
0.11 град./сек.2
(Clark, Stewart, 1969).
Говоря
о порогах чувствительности полукружных
каналов, необходимо указать, что
приведенные выше величины получены в
результате регистрации нистагма, т. е.
процесса, уже «реализованного»
вестибуломоторной «рефлекторной
дугой». Если же рассматривать феномены
в сенсорной части такой дуги, то
представление о чувствительности
вестибулярных рецепторов совершенно
меняется. Показано, например, что кривые
чувствительности полукружных каналов
(по изменению постоянных потенциалов
купулы) проходят через ноль системы
координат, где по оси абсцисс отложена
сила стимула, а по оси ординат — величина
ответа (Trincker, 1957). На графике практически
невозможно найти наименьший угол
отклонения купулы, при котором реакция
отсутствует. Аналогичные данные
приводят и другие авторы (de Vries, 1948; de
Vries, Schierbeek, 1953). Это не должно казаться
странным, потому что имеются сведения,
указывающие на чрезвычайно высокую
чувствительность и других (а не только
вестибулярных) рецепторов. Можно
указать на вычисления Бекеши, касающиеся
величины движений базилярной мембраны
кортиева органа, на уровне порога не
превышающих диаметра атома водорода
(цит. по: Dohlman, 1959). Органы боковой линии
некоторых рыб могут реагировать на
перемещения порядка 1 А (цит. по: Bar- low,
1964). Высокая чувствительность рецепторных
систем является общим правилом, и
вестибулярные рецепторы в этом смысле
не представляют исключения.
Отклонение
купулы может быть вызвано не только
угловым ускорением. Эксперименты
Эвальда (Ewald, 1892) и их более поздние
модификации показали, что отклонения
купулы можно получить с помощью
создаваемого гидромеханически давления
(положительного или отрицательного).
С помощью этой мето
69
дики
была найдена зависимость угла отклонения
купулы и линейного смещения эндолимфы
в полукружном канале (Dohlman,. 1935), а также
связь между углом отклонения купулы и
электрическими изменениями в
полукружных каналах (Trincker, 1957),. Ее
применение позволило выявить связь
отклонений купулы; с движениями глаз
(Steinhausen, 1933; Vilstrup, 1950) и исследо-- вать
морфо-физиологические корреляции между
отдельными; каналами и изолированными
экстраокулярными мышцами (Lorente* de Nd,
1933; Szentagothai, 1950, 1952). Несмотря на определенные*
достоинства такого способа стимуляции
(прежде всего его относительная
локальность), следует иметь в виду
связанные с ним серьезные технические
трудности, малую надежность эксперимента
и ту опасность, которую введение в канал
микроканюли диаметром 50 мкм
представляет для сохранения нормального
уровня; лабиринтной афферентной
импульсации. Кроме того, как показывают
данные Спёндлина (Spoendlin, 1966), введение
физиологического раствора в эндолимфу
латерального канала на 15 мин., вызывает
специфические патологические изменения.
В основном? они затрагивают опорные
клетки crista ampullaris и, реже, сенсорные
клетки, которые могут реагировать на
инъекцию сильным; выпячив анием пр
отопл азмы.
В
физиологическом эксперименте и в
клинике распространена? температурная
стимуляция лабиринта, эффект которой
впервые- правильно объяснил А. Борнгардт
(1875). В 1906 г. Роберт Барани сообщил о
нистагме, возникающем при температурной
стимуляции лабиринта теплой или холодной
водой. Такой способ исследования?
вестибулярного аппарата он назвал
калорическим (Вагапу, 1906).. Барани
разработал метод клинических проб и
рассмотрел физические явления в
полукружных каналах при калоризации.
Причиной появления калорического
нистагма Барани считал конвекционные
токи эндолимфы, возникающие в результате
изменения ее плотности при нагреве или
охлаждении. При этом величина реакции
зависит от разницы температур тела и
воды, а также от ориентации канала. При
одной и той же температуре максимум
реакции соответствует вертикальной
ориентации плоскости канала. При
сравнении действий различных температур
необходимым условием является одинаковая
ориентация канала.6
Позднее
Кобрак (Kobrak, 1918) модифицировал калорическую
пробу Барани, уменьшив количество воды,
орошающей ухо, с 200—250 до 5 мл. Способы
калорической стимуляции лабиринта в
клинике (орошение водой наружного
слухового прохода) со вре-
6 Количественное
описание феномена калорической
стимуляции полукружного канала
человека с учетом физических свойств
эндолимфы (вязкость, плотность,
температурный коэффициент вязкости и
коэффициент температурного расширения)
дается в работе (Steer et al., 1968).
70
йен
Барани изменились мало? Они не свободны
от недостатков, сводящихся в основном
к трудностям дозирования и невозможности
строгой локализации раздражителя. С
точки зрения клиницистов основным
преимуществом калорической пробы
является возможность раздражения
одного лабиринта. Однако следует иметь
в виду, что это справедливо только для
обнаружения далеко зашедшей патологии
(выключение лабиринта). Судить же об
асимметрии лабиринтной функции на
основе только калорической пробы
следует с осторожностью (см. стр. 81:
Sala, 1965). Изменения, внесенные за
последнее время в методику калорических
тестов, сводятся к достаточно точному
(до 0.1° С) термостатированию воды,
используемой для орошения (Dohlman, 1961, и
др.).
Известно,
что эндолимфатические токи возникают
в полукружных каналах даже при
разнице температур в 0.5° (Maier, Lion, 1921).
Для стеклянных капилляров, имитирующих
каналы, эта величина составляет 0.1°
(Ундриц, 1927). Результаты вычислений
(Schmaltz, Volger, 1924) показывают, что скорость
тока эндолимфы, вызывающего нистагм
при «минимальном орошении», не превышает
величины 2х10~5
см/сек., а нистагм начинается тогда,
когда смещение эндолимфы составляет
всего 1 мкм.
При
классической калорической пробе
вестибулярная реакция во многом зависит
от так называемых экстралабиринтных
факторов, т. е. от толщины тканей,
через которые происходит теплообмен
между водой и эндолимфой, конфигурации
наружного слухового прохода,
особенностей строения височной кости,
а также сосудистого и нервного компонентов
реакции (Ino, Okamoto, 1963).
Методика
локального (точечного) нагревания
обнаженных полукружных каналов в
остром эксперименте сводит до минимума
роль экстралабиринтных факторов,
позволяет вызвать у животных (голуби)
калорический нистагм и определить его
температурный порог. Для этого
используется медленное управляемое
нарастание температуры и ее регистрация
микротермистором (Богданов, Орлов,
1965).
Прямое
нагревание отдельных полукружных
каналов (без записи температуры) в
сочетании с экстраклеточной регистрацией
активности нейронов глазодвигательных
ядер позволило выделить в этих ядрах
единицы, реагирующие на стимуляцию
горизонтального и верхнего каналов
(Manni, Desole, 1966).
В
настоящее время в клинике широко
используется калорическая проба по
методу Фитцжералда и Холпайка
(Fitzgerald, Hallpike, 1942), которая заключается
в орошении наружного слухового прохода
водой с температурой 30 и 44° С, т. е. на
7° ниже и выше температуры тела. При
этом время орошения составляет 40
сек., а объем воды —* 400 мл для каждого
уха. Метод позволяет выявить
патологию, выражающуюся в вестибулярной
асимметрии (эта асимметрия может иметь
как периферическую, так и центральную
природу).
71
Перечисленные
способы стимуляции полукружных каналбв
связаны с функцией купулярного аппарата.
Это подразумевает применение тестов,
вызывающих появление более или менее
длительных токов эндолимфы и отклонения
купулы. Вторая группа методик не связана
с механическими изменениями в купуло-
эндолимфатической системе. Сюда
относится прежде всего электростимуляция
вестибулярного аппарата в целом или
отдельных его частей, в том числе
полукружных каналов. Гальвано-вестибулярные
реакции известны давно (см. обзоры по
этому вопросу: Spiegel, Sommer, 1944а; Хечинашвили,
1958). Поляризация лабиринтов постоянным
током дала возможность исследовать
изменения афферентной импульсации
в вестибулярном нерве (Lowenstein, 1955) и
связанные с этим реакции нейронов в
различных отделах ЦНС (de Vito et al., 1956;
Arduini, Pompeiano, 1957; Grosser et al., 1959; Pompeiano, Cotti,
1959; Горгиладзе, 1964a; Лещинюк, 1968a, и др.),
а также вестибуло-спинальные эффекты
(Spiegel, Sommer, 1944a и др.). В отношении механизма
действия постоянного тока на вестибулярный
аппарат известно, что гальванизация
полукружного канала не вызывает
отклонений купулы (St einhausen, 1933).
Использование
одиночных электрораздражений
вестибулярного нерва в сочетании с
отведением вызванных потенциалов
позволило локализовать вестибулярную
проекционную зону в коре больших
полушарий и других отделах ЦНС (Andersson,
Gernandt, 1954; Mickle, Ades, 1954).
Другой
вид электрораздражения вестибулярного
аппарата (нерва или рецептора) —
ритмическая стимуляция электрическими
импульсами, имитирующими вестибулярную
афферентную импуль- сацию, дал возможность
исследовать вестибулоглазные (Dumont-
Tye, Dell, 1961) и вестибулоспинальные (Орлов,
1962а) рефлекторные дуги, а также роль
вестибулярной афферентной импульсации
в формировании нистагма и тонических
отклонений глаза (Левашов, 1965а, 19656).
Существуют
способы стимуляции, основанные на
микрофонном эффекте волосковых
клеток вестибулярного сенсорного
эпителия различных животных. Это
так называемый «эффект Туллио», впервые
продемонстрированный на голубях
(Tullio, de Marco, 1938). С помощью стимуляции
звуковыми и ультразвуковыми колебаниями
в диапазоне от 0.3 до 100 кгц показан
микрофонный эффект любого из полукружных
каналов, саккулюса и утрику- люса морской
свинки с полностью разрушенной улиткой
(Trin- cker, Partsch, 1959). Микрофонные свойства
и высокая чувствительность
вестибулярного эпителия обусловливают
появление в шейных мышцах моносинаптических
потенциалов при кратковременных
раздражениях перепончатой ампулы
горизонтального канала голубя толчками
пьезокристалла (Орлов, 19626). Сходные эф
72
фекты
наблюдаются и при действии звуковых
щелчков (Eyck, 1950, и др.).
В
последнее время появились данные о
действии биологически активных веществ,
введенных в полукружные каналы. Инъекции
ацетилхолина, антихолинэргичёских
веществ (атропин, тетраэтил аммоний)
, ингибитора холинэстеразы (диизопропил
флю- орофосфат) в горизонтальный канал
кролика вызывают различные изменения
вестибулярного нистагма. Однако, как
указывают авторы, в этом случае нельзя
точно определить количество веществ,
действующих на нейроэпителий. Кроме
того, наряду с их влиянием на передачу
импульсов в месте контакта рецептора
с нервным волокном нельзя исключить и
прямой фармакодинамический эффект
(Rossi et al., 1964).
Функция
отолитовых рецепторов. Способы стимуляции
и пороги чувствительности отолитовых
органов. Известно, что отолитовые
органы способны функционировать как
«рецепторы положения», т. е. изменять
частоту афферентной импульсации в
вестибулярном нерве при изменении
положения головы относительно
направления силы гравитации (Ross, 1936, и
др.). Можно считать, что в настоящее
время на равных правах сосуществуют
две группы фактов, по-разному объясняющих
механизм отолитовой функции. Одна
из них подтверждает «теорию скольжения»,
впервые в общем виде сформулированную
Брейером (Breuer, 1891). Согласно этой
концепции, раздражающим фактором
здесь является скольжение отолитовой
мембраны, в результате которого
происходит тангенциальное сгибание
волосков сенсорных клеток. Прямые
эксперименты подтверждают правильность
такой точки зрения. С помощью рентгеновских
лучей показано (de Vries, 1950), что отолитовая
мембрана саккуляр- ной макулы отклоняется
тангенциально при любых центростремительных
ускорениях, вплоть до 11 g, когда величина
её смещения составляет 0.23 мм. Аналогичные
данные приводит Виль- струп (Vilstrup, 1951):
при крайних наклонах тела отолит
сдвигается тангенциально на 15 мкм
в любую сторону, в зависимости от
направления наклона тела. Эту точку
зрения энергично отстаивает Тринкер
(Trincker, 1962), который указывает, что только
тангенциальные смещения отолитовой
мембраны утрикулюса вызывают
биоэлектрические эффекты. Это совпадает
и с более ранними результатами,
полученными при использовании других
методик (Trincker, 1962).
Однако
вторая группа фактов подкрепляет старые
теории, согласно которым максимум
возбуждения макул возникает при
отвисании отолитовой мембраны и,
следовательно, при натяжении волосковых
клеток (Magnus, 1924), либо при давлении
отолитовой мембраны на волосковые
клетки (Quix, 1925). Такие данные получены
в последнее время японскими исследователями
и позволяют считать, что и утрикулярная,
и саккулярная макулы чув
73
ствительны
к ускорению, действующему в вертикальном
направлении. При этом система
реагирует как на давление, так и на
натяжение волосков, хотя более эффективным
является давление. Обе макулы обнаруживают
значительную степень дублирования
функций, работают совместно и чувствительны
к линейным ускорениям любого
направления (Owada, Shiizu, 1960; Sasakiet al., 1963;
Sasaki, 1967).
Резюмируя,
можно сказать, что ни механизм скольжения,
с одной стороны, ни механизм натяжения
или давления — с другой, не могут в
отдельности объяснить всего многообразия
фактов, накопленных в физиологии
отолитовых органов.1
Вероятно,
трудности и противоречия в трактовке
отолитовых механизмов являются
результатом недооценки сложности
картины: особенностей деформации
отолитовой мембраны, ориентации
волосковых клеток и функциональной
локализации в макулах. Говоря об
отолитовой мембране, следует отметить,
что возможны деформация этой мембраны
и смещение ее в любом направлении (к
макуле, от макулы или тангенциально).
В каждом случае деформация волосковых
клеток различна (соответственно сжатие,
чтатяжение, отклонение). Ситуация
осложняется еще и наличием в макулах
функциональной локализации, т. е.
функциональной связи разных отделов
макулы с различными мышечными группами.
Успешную попытку рассмотрения функции
макул с учетом перечисленных выше
особенностей предпринял Сентаготаи
(Szentagothai, 1952) на основе анализа рефлексов
глазных и соматических мышц при
дифференциальной стимуляции отолитового
аппарата.8
9
По
современным представлениям, в отличие
от купулы, которая по своим механическим
свойствам является сверхкритически
демпфированным торсионным маятником,
отолитовая мембрана представляет собой
гармонический осциллятор (de Vries, 1956).
Адекватным стимулом для отолитовых
рецепторов являются линейные
ускорения или изменения положения
отолита в гравитационном поле.
Линейные ускорения могут быть получены
в условиях
8 Многие
анатомические вариации отолитового
аппарата, в частности кривизна макул
и пространственное расположение их
передних краев, гармонируют с образом
жизни и в особенности с локомоцией
каждого данного вида позвоночных. У
птиц, подвергающихся действию вертикальных
ускорений, утрикулярная макула
велика, а саккулярная — мала; у водных
животных картина обратная.
Сравнительноанатомические исследования,
в которых прослежена эта связь
(Mygind, 1969), показали также существование
дифференциальной иннервации макул
(в особенности утрикулярной), в которых
выделены части с самыми высокими
волосковыми клетками («доминирующие»
части, иннервируемые более мощными
периферическими волокнами отолитового
нерва), и противоположные части
(«подчиненные», снабжаемые менее мощным
пучком волокон из центральной части
нерва). Между ними может находиться
промежуточная «слепая» зона, лишенная
иннервации.
9 Теория
таких маятников в доступной форме
изложена в переведенной на русский
язык книге Ф. Гродинза (Grodins, 1963).
74
экспёрймёнта
с помощью параллельных качелей (Хилов,
1933) либо при прямолинейном движении по
земле или в воздухе. Данные об
исследовании функции отолитовых органов
с помощью параллельных качелей в
сочетании с методом электронистагмогра-
фии подробно описаны в обзорной работе
(Jongkees, Philipszoon, 1964). Различные величины
центростремительных ускорений,
взаимодействующих с ускорением
гравитации и смещающих отолиты в
направлении равнодействующей, в
лабораторных условиях можно получить,
вращая объект на центрифуге и создавая
ситуации, соответствующие «повышенной»
или «пониженной» весомости (например,
для исследования вестибулярной функции
в условиях имитации авиационных и
космических полетов).10
Литературные
данные указывают на высокую чувствительность
лабиринта к центростремительным и
линейным ускорениям. Отчетливые
сдвиги биоэлектрической активности в
мозжечке кошки (флоккулонодулярная
доля) наблюдаются при ускорениях,
равных 0.005—0.01 g (Margaria, 1958). Дифференциальный
порог по линейному ускорению (на
центрифуге) составляет 0.0003 g, а порог
чувствительности к линейному ускорению
(в самолете) — около 0.07 g (цит. по: Clark,
1963). По данным, полученным в результате
регистрации электрической активности
«отолитовых» нейронов бульбарного
комплекса интактной лягушки (в условиях
нормальной гравитации), порог линейного
ускорения меньше 1 см/сек.2
при продолжительности стимула менее
5 мсек. (Gualtierotti, 1967).
Измерения,
проведенные на людях с помощью
высокочувствительных акцелерометров,
показали, что сердечные удары вызывают
линейные смещения головые ускорением
порядка 0.01 — 0.02 g. У человека удавалось
зарегистрировать линейные ускорения
головы, которые в 10 раз превышают порог
чувствительности отолитовых органов
(Gualtierotti, 1967, 1968). Избежать этого удавалось
только с помощью самой жесткой фиксации
головы. Высказана гипотеза, согласно
которой сердечные удары обусловливают
постоянный тонус отолитовых рецепторов.
Так или иначе, под влиянием таких
ускорений активность отолитовых
рецепторов лягушки уже может быть
модифицирована (Gualtierotti, 1968). Это лишний
раз подтверждает строгость требований,
предъявляемых к экспериментам на
вращающихся стендах, где к контролируемым
ускорениям неизбежно присоединяется
неконтролируемая вибрация, создаваемая
работой двигателя и трущихся частей.
Структура
и функция отолитовых рецепторов
исследуются в последнее время в
аспекте условий авиационных и космических
по
10 Предложено
большое количество методик для стимуляции
отолитов и полукружных каналов, а также
ряд функциональных тестов для оценки
вестибулярной функции (см.: Воячек,
1927; Стрелец, 1962а, 19626).
75
летов
со всем разнообразием возникающих при
этом факторов: ускорениями, перегрузками,
невесомостью.11
Показано, что в волосковых клетках
и синапсах утрикулюса под влиянием
центростремительного ускорения (10
g в течение 3 мин.) наблюдаются
цитохимические сдвиги, связанные с
функцией нуклеиновых кислот (Винников,
1964). Что касается морфологических
изменений в сенсорном эпителии
макул, то электронномикроскопические
исследования указывают на их
отсутствие при интенсивных перегрузках
(5.5 или 11 g в течение 10 мин.) (Spoendlinet al.,
1964). Другим важным фактором авиационных
и космических полетов, почти не
встречающимся в нормальных условиях,
является невесомость. Регистрация
микроэлектродом активности вестибулярного
нерва у кошки во время кратковременного
(1.2 сек.) свободного падения показала
резкое увеличение частоты разрядов
(Fiorica et al., 1962). Однако кратковременность
состояния невесомости не позволила
проанализировать ее влияние на
чувствительность отолитового аппарата
к линейным ускорениям. Данный вопрос
исследовался с помощью регистрации
активности одиночных волокон
вестибулярного нерва лягушки,
связанных с отолитовыми рецепторами.
В состоянии невесомости, вызванном
полетом самолета по кепле- ровской
кривой, вначале наблюдалось увеличение
ответов на линейное ускорение, а
после примерно 10-секундного полета с
нулевой гравитацией — внезапное
блокирование ответа на линейное
ускорение. Авторы считают, что это
происходит благодаря механизму,
предохраняющему центры от слишком
интенсивного афферентного потока.
Отмечалось также резкое усиление общей
активности вестибулярного нерва в
результате серии кратковременных
параболических полетов. Эффекты
невесомости расцениваются как результат
дефицита раздражения отолитового
аппарата по аналогии с затемненной
сетчаткой (Gualtierotti, Gerathewohl, 1965).
Вестибулярная
афферентная импульсация и ее изменения
под влиянием стимуляции полукружных
каналов и отолитовых органов. Во время
ускоренного вращения возбуждаются в
основном каналы, плоскость которых
соответствует плоскости приложенного
ускорения. Однако имеются отдельные
указания на то, что горизонтальные
каналы реагируют на вращение лишь
вокруг вертикальной, основной оси,
не возбуждаясь при поворотах вокруг
любой из двух горизонтальных осей,
между тем как вертикальные каналы
отвечают на вращение вокруг всех трех
осей (Lowenstein, Sand, 1940а). Впрочем это было
показано только на хрящевых рыбах;
в отношении теплокровных животных
таких сведений нет. Не исключено, что
при вращении вокруг любой оси стимулируется
не только «основная» пара каналов, но
(отчасти) и другие каналы.
Исследования,
в которых регистрировалась афферентная
импульсация в отдельных волокнах
вестибулярного нерва в покое
11 Анализ
этих факторов см.: Комендантов, 1965.
76
й
прй адекватной стимуляции, показали,
что ампулопетальныё
токи эндолимфы
(и следовательно, утрикулопетальные
отклонения
купулы) в горизонтальных
полукружных каналах и ампулофу-
£альные
токи — в вертикальных каналах вызывают
увеличение
частоты афферентной
импульсации. При противоположно
направ-
ленных токах эндолимфы
наблюдался обратный эффект (Ross,
1936;
Lowenstein, Sand, 1940а, 1940b, и др.). Из этого следует,
что
рецепторы кристы характеризуются
бидирекционной, симметрич-
ной
чувствительностью.
Специальное
изучение во-
проса показало, что в
оп-
ределенном (так называе-
мом
физиологическом) ди-
апазоне
положительных и
отрицательных
ускорений
имеет место
одинаковая
чувствительность
рецепто-
ров полукружных каналов
к
стимулам обоих направ-
лений (при
условии их
физического равенства).
При
этом в нервных ве-
точках полукружных
ка-
налов регистрируются рав-
ные
по величине (по отно-
шению к уровню
спонтан-
ной активности) и
проти-
воположные по знаку
сдвиги
частоты импульсов
(Groen et al., 1952;
Groen,
1961b). Аналогичные ре-
Рис.
28. Различные типы реакции
(А, В, С, D)
изолированных волокон ве-
стибулярного
нерва ската (Groen, 1961b).
По
оси абсцисс —
скорость вращения перед
«стоп-стимулом»
в град./сек.; по
оси ординат —
количество
спайков в 1 сек. Плюсом
обозначен
«стоп-стимул», вызывающий
утрикулопетальное,
минусом
—
утрикулофугальное отклонение ку-
пулы.
зультаты
получены и в тех
случаях,
когда в качестве показателя использовалась
не афферентная импульсация, а какая-либо
из моторных реакций — движения глаз
(нистагм) (Hallpike, 1961; Hood, 1961) или повороты
головы у животных, которые не обнаруживают
реакции вестибулярного нистагма
(лягушки) (Gribenski, 1963).
Приведенные
факты послужили основанием для пересмотра
одного из положений физиологии
полукружных каналов, получившего в
научной литературе название «второго
закона Эвальда». Согласно данным опытов
Эвальда (Ewald, 1892), рецепторы полукружных
каналов обладают неодинаковой
чувствительностью к токам эндолимфы
разных направлений: в горизонтальном
канале ампулопетальный ток эндолимфы
вызывает более сильную реакцию, чем
ампулофугальный; в вертикальных каналах
имеют место обратные соотношения. Эти
данные были получены при сильных
(форсированных) сдвигах эндолимфы в
каналах.
77
Новые
йсследования показалй, что в физиологическом
диапазоне раздражений сенсорные
области полукружных каналов интактного
лабиринта с большой точностью дают
равные по величине и противоположные
по знаку ответы на движение эндолимфы
в обоих направлениях. Вне этого диапазона
первоначальное заключение Эвальда
остается в силе (Groen, 1961b).
В
нервах полукружных каналов имеются
различные группы волокон, реакции
которых перекрывают физиологический
диапазон стимулов, как положительных,
так и отрицательных. Результаты,
полученные при исследовании лабиринтной
импульсации ската, позволили выделить
4 типа волокон (рис. 28): А, В, С и D (Groen et
al., 1952; Groen, 1961b). Волокна типов А и В, так
называемые пропорциональные волокна
(количественно преобладающие группы),
характеризуются бидирекционной
чувствительностью. Они активируются
утрикулонетальным (+) и тормозятся
утрикуло- фугальным (—) отклонениями
купулы. Типы А и В различаются по крутизне
характеристики, чувствительности и
максимальной частоте ответов. Эксперимент,
в котором регистрировалась активность
волокон типов А и В (изолированный
лабиринт ската, горизонтальный
полукружный канал), показал, что ответ
таких волокон, выраженный в количестве
потенциалов действия за секунду как
функции стимула, имел вид S-образной
кривой, подобной анодно-сеточной
характеристике электронной лампы-триода.
Линейная часть этой характеристики
находилась в диапазоне стимулов от
+40 до —40 град./сек.12
Волокна третьего типа (С) не реагировали
изменением частоты разрядов на
положительные стимулы. Для подавления
их активности требуется сильный (до
100 град./сек.) отрицательный стимул.
Наконец, у волокон типа D (этот тип,
как и тип С, встречается относительно
редко) спонтанная активность отсутствует.
Импульсация в этих волокнах появляется
под влиянием сильных положительных
стимулов (от 100 град./сек. и выше).
Волокна типов С и D можно считать
связанными с монодирекционными
сенсорными элементами. Исследования
на теплокровных (кошки) позволили
выделить 3 типа ампулярных волокон
(Gernandt, 1949). Тип I Гернандта^. (количественно
основной, 83% единиц) соответствует
группам А и В Гроенаи соавт. Тип II (12%
единиц) усиливал активность под влиянием
как положительных, так и отрицательных
стимулов. Тип III (5% единиц) тормозился
при действии стимула любого знака.
12 Адекватным
раздражителем для полукружного канала
является угловое ускорение, в связи
с чем правильнее измерять стимул в
величинах углового ускорения, т. е.
в град./сек.2
или, согласно «Международной системе
единиц» (СИ), в рад./сек.2
Однако авторы большинства опубликованных
работ до настоящего времени,
характеризуя стимул, приводят величины
угловой скорости. При этом имеют в
виду конечную скорость, достигнутую
при разгоне установки или перед внезапной
остановкой вращения. При стоп- стимуле
(внезапной остановке) указанием на
длительность торможения стенда обычно
неоправданно пренебрегают^
78
Поскольку
волокна типа I (Gernandt, 1949), или типов А и
В (Groen, 1961b), количественно преобладают,
можно предположить, что именно они
определяют верхнюю границу физиологического
диапазона стимулов. По-видимому,
минимальный стимул, вызывающий
снижение спонтанной импульсации до
нуля при ампу- лофугальном отклонении
купулы в горизонтальном канале,
удваивает импульсацию при
ампулопетальном ее отклонении. Вполне
вероятно, что верхней границей
физиологического диапазона для единиц
типа А и В можно считать стимул,
результатом которого является удвоение
частоты импульсации по сравнению с
уровнем покоя. На основании исследований
нистагма можно считать, что эта граница
соответствует стоп-стимулу при вращении
со скоростью 60—70 град./сек. (Groen, 1961b;
Бохов, Шипов, 1965).
Основные
сведения о реакциях отолитовых рецепторов
на адекватные стимулы были получены
в исследованиях, проведенных на хрящевых
рыбах (Lowenstein, Roberts, 1949). В афферентных
нервных веточках, иннервирующих
отолитовые рецепторы, во всех случаях
регистрируется спонтанная активность.
На основании измерений частоты
импульсации Лёвенштейн и Робертс
разделяют отолитовые рецепторы на
медленно адаптирующиеся статические
(т. е. собственно «статорецепторы» —
рецепторы положения) и быстро
адаптирующиеся динамические, реагирующие
на изменения положения головы с
постоянной скоростью («рецепторы
неправильного положения»). Электрическая
активность, соответствующая первому
типу рецепторов, изменялась при
отклонении от нормали и достигала
максимума в положении головы «носом
вверх» (повороты в сагиттальной
плоскости). В случае поворотов головы
во фронтальной плоскости максимум
соответствовал положению, при котором
исследуемый лабиринт находился вверху.
Когда препарат фиксировали в новом
положении, частота в течение минут
оставалась резко отличной от нормальной.
Вторая группа рецепторов обнаружила
значительное падение активности при
выведении препарата из нормального
положения, независимо от направления
движения. Если последнее прерывалось,
то активность возвращалась к норме
независимо от положения головы в точке
остановки. Такие рецепторы не могут
считаться «статорецепторами», поскольку
они сигнализируют о перемещении головы
в пространстве. Разница между
«рецепторами неправильного положения»
и рецепторами полукружных каналов
состоит в том, что первые отвечают
на наклоны головы с постоянной скоростью,
тогда как вторые реагируют на угловое
ускорение.13
Отолитовые органы реагируют на
линейные перемещения во всех трех
координатах. По данным Лёвенштейна и
Робертса, рецепторы саккулюса фактически
не отличаются от рецепторов утрикулюса
по своей способ
13 Аналогичные
типы реакций были получены на нейронах
бульбарного комплекса костистых рыб
(Schoen, 1957). Кроме того, там встречались
нейроны, отвечавшие активацией на
боковые наклоны в обе стороны.
79
ности
сигнализировать о наклонах головы. На
основании осциллографической
регистрации авторы установили, что в
передних двух третях саккулярной макулы
рецепторы вибрации преобладают над
рецепторами гравитации, тогда как в
задней области имеют место обратные
отношения.
У
лягушки (Gualtierotti, Gerathewohl, 1965) были обнаружены
гравитационные рецепторы, отвечающие
на наклоны головы увеличением частоты
разрядов (логарифмическая зависимость
в соответствии с законом Вебера—Фехнера).
Частота разрядов той же единицы снижалась
при наклоне в противоположную сторону,
однако логарифмическая зависимость в
этом случае не наблюдалась.
Почти
полное отсутствие в литературе сведений
о реакциях рецепторов преддверия
млекопитающих (по электрофизиологическим
показателям) объясняется серьезными
техническими трудностями, с которыми
сопряжена регистрация электрической
активности нерва преддверия у этих
животных. Данные о том, что нейроны
ганглия Скарпа кошки, связанные с
отолитовыми рецепторами, не
обнаруживают непосредственных сдвигов
частоты им- пульсации при любом изменении
положения головы (такие сдвиги если и
наблюдаются, то лишь спустя примерно
40 сек. — Wing, 1963), совершенно не поддаются
объяснению с позиций общеизвестных
«отолитовых» концепций и, кроме того,
не согласуются с данными, полученными
при регистрации активности нейронов
вестибулярных ядер (Adrian, 1943, и др.).
Эфферентные
влияния. В настоящее время существует
представление об эфферентной
иннервации вестибулярного аппарата
(см.: Brodal el al., 1962). Методом электронной
микроскопии показана двойная
иннервация вестибулярного эпителия и
обнаружены как постсинаптические, так
и пресинаптические структуры (Wer- sall,
1958; Engstrom, 1958, 1961; Engstrom, Wersall, 1958). Количество
эфферентных вестибулярных волокон
относительно мало (около 400) в сравнении
с общим числом волокон в вестибулярном
нерве, составляющем у кошки 12000 (Gacek et
al., 1965). Диаметр большинства эфферентов
у кошки равен 2—4 мкм, т. е. по толщине
они соответствуют афферентам (Gacek,
1960).
Происхождение
вестибулярных эфферентов в настоящее
время выяснено лишь частично. Установлено,
что после перерезки вестибулярного
корешка (перекрещенный оливарный пучок)
эфферентные волокна, обнаруживаемые
в норме в утрикулюсе, саккулюсе и кристе,
перерождаются (Petroff, 1955). Позднее на
морских свинках было показано
существование «прямого вентрального
вестибулярного эфферентного пучка»,
возникающего в неизвестном ранее
«вставочном вестибулярном ядре»,
расположенном вентральнее ядра Дейтерса
и дорсальнее нисходящего ядра. Обнаружен
и «прямой дорсальный эфферентный
вестибулярный пучок», идущий из переднего
отдела каудальной части ядра Дейтерса.
Оба пучка
80
являются
частью эфферентной вестибуло-кохлеарной
системы (Rossi, Gortesina, 1962).
Имеются
также данные об эфферентной регуляции
вестибулярных рецепторов.
Гистохимические методы позволили
обнаружить в вестибулярном сенсорном
эпителии холинэстеразу (Dohlman et al., 1958;
Dohlman, 1960b; Ireland, Farkashidy, 1961), которая
обычно локализуется в эфферентных
окончаниях. Последние найдены на клетках
типа II Вёрселла (Wersall, 1956; Engstrom, 1958). С
помощью электрофизиологических методов
было зарегистрировано появление
эфферентной импульсации в центральных
концах нервов, отделенных от лабиринта,
при стимуляции полукружных каналов
(Schmidt, 1963; Bertrand, Veenhof, 1944; Gleisner, Henriksson, 1964)
или отолитовых органов другой стороны
(Bertrand, Veenhof, 1964). Наряду с этим показано,
что электростимуляция эфферентной
вестибулярной системы (ЭВС) в области
ядра Дейтерса всегда вызывает
гиперполяризационные сдвиги постоянных
потенциалов в обоих лабиринтах (Sala,
1965), что соответствует тормозному
эффекту (Lowenstein, 1955; Trincker, 1957). При этом
модифицируется и афферентная импульсация
в вестибулярном нерве. Имеются прямые
указания на существование петли обратной
связи между лабиринтами; теплая вода
вызывает деполяризацию в ипсилатеральном
и гиперполяризацию — в контрлатеральном
лабиринте, холодная вода оказывает
противоположное действие (Sala, 1965).14
Таким
образом, можно считать доказанным
наличие тормозящего и регулирующего
действия ЭВС на чувствительность
вестибулярных рецепторов, хотя
количественная сторона проблемы не
разработана.
ВЕСТИБУЛЯРНЫЕ
ЯДРА ПРОДОЛГОВАТОГО МОЗГА
Первичные
вестибулярные волокна, берущие начало
в ганглии Скарна, оканчиваются на
нейронах вестибулярных ядер продолговатого
мозга своей стороны. В продолговатом
мозгу различают четыре основных
ядра, относящихся к вестибулярной
системе: верхнее вестибулярное ядро
(ангулярное, или ядро Бехтерева),
латеральное вестибулярное ядро
(Дейтерса), медиальное вестибулярное
ядро (дорсальное, главное, триангулярное,
или ядро Швальбе) и нисходящее
вестибулярное ядро, которое называют
также спинальным, или нижним ядром (см.
классификацию: Brodal et al., 1962). Вопрос о
связях между вестибулярными ядрами
правой и левой половин продолговатого
мозга изучен недостаточно. Высказывались
предположения, что эти связи осуществляются
волокнами (или их коллатералями),
отходящими от нерв
14 В
этой работе, выполненной на кошках,
регистрировались потенциалы в области
crus communae; активный электрод находился
на эндостеуме или в перилимфе.
(э
Сенсорные системы
81
ных
клеток ядер одной стороны и оканчивающимися
на клетках ядер другой стороны (Brodal et
al., 1962). С такой точкой зрения не совпадали
электрофизиологические данные,
показывающие, что при электростимуляции
вестибулярного нерва вызванные ответы
возникают только в ядрах на стороне
раздражения. На противоположной
стороне ответы отсутствовали (Mickle,
Ades, 1954, и др.). Другие авторы на основании
микроэлектродных исследований
полагали, что взаимные связи между
ядрами двух сторон являются опосредованными
и осуществляются через ретикулярную
формацию ствола мозга (Gernandt, Thulin, 1952).
Некоторая ясность в этот вопрос была
внесена работой Шимазу и Прехта (Shi-
mazu, Precht, 1966), которые провели исследования
на децеребрированных кошках без
применения наркотиков. Они отводили
стеклянными микроэлектродами фокальные
потенциалы (field potentials) от различных
участков вестибулярных ядер при
электростимуляции контрлатерального
вестибулярного нерва. Зоны ответов
были локализованы в вентральной части
медиального ядер, а также в вентро-медиальной
части верхнего и латерального ядра.
Разрез мозгового вещества по передней
линии дна IV желудочка на глубину 2 мм
(от нижнего двухолмия до obex) уменьшал
или полностью снимал начальный позитивный
и последующий медленный негативный
компоненты ответов контрлатеральных
вестибулярных ядер, что, по мнению
авторов, связано с перерезкой
комиссуральных волокон. Полифазные
ответы, вызываемые (после перерезки)
более сильной стимуляцией вестибулярного
нерва, авторы рассматривают как
результат прохождения импульсов через
нейроны ретикулярной формации.
Физиологические данные о комиссуральных
связях между вестибулярными ядрами
двух сторон нуждаются в морфологическом
подтверждении.15
Вестибулярные
ядра являются первым уровнем в центральной
нервной системе, где происходит обработка
информации о движении или изменении
положения тела в пространстве, поступающей
от рецепторов лабиринта. Впервые
исследование активности одиночных
вестибулярных элементов продолговатого
мозга кошки было осуществлено
Эдрианом (Adrian, 1943). Им были обнаружены
единицы, реагирующие изменением
частоты импульсов на раздражение
отолитовых органов. Принадлежность
единиц к определенным ядрам не
определялась. Стимуляция отолитовых
органов (наклоны животного) приводила
к увеличению частоты разрядов, если
наклон производили в ипсилатеральную
сторону. При наклоне в контрлатеральную
сторону импульсная активность
затормаживалась. Были исследованы
также единицы, реагирующие на линейные
ускорения и наклоны в других плоскостях.
Некоторые единицы сохраняли высокий
уровень импульсации в течение всего
15 Краткую
историческую справку по этому вопросу
см. в работе Г. И. Горгиладзе (19666),
времени
длительно поддерживаемого наклона
животного. В работе Руперта и соавт.
(Rupert et al., 1962), подтвердивших данные
Эдриана, наряду с единицами, отвечавшими
на медленные наклоны головы и на
измененное статическое положение
головы в пространстве, были выделены
единицы, у которых продолжительность
межспайковых интервалов изменялась
только в момент быстрых наклонов. В
латеральном ядре большинство клеток
реагирует на наклоны вокруг продольной
оси тела. Каждый лабиринт вызывает
эффекты как в ядре своей, так и в ядре
противоположной стороны (Fujita et al., 1968).
Описанные
реакции Эдриан связывал с функцией
рецепторов саккулюса, опираясь на
теорию Магнуса (Magnus, 1924). Однако необходимых
доказательств для этого утверждения
в работе Эдриана не содержится.
Описанные им эффекты могли быть связаны
не только с натяжением или сдавливанием
волосков отолитом саккулюса, но и с
деформацией волосков утрикулюса
вследствие тангенциальных смещений
утрикулярного отолита при наклонах.
При
действии угловых ускорений изменения
импульсной активности вестибулярных
единиц, описанные Эдрианом, можно было
сопоставить со сдвигом в
купулоэндолимфатической системе
полукружных каналов. Единицы,
реагировавшие на раздражение
горизонтальных каналов, увеличивали
частоту импульсов при вращении в
ипсилатеральную сторону, что
соответствовало ампу- лопетальному
сдвигу эндолимфы и купулы в канале на
стороне отведения. При вращении в
контрлатеральную сторону, когда
эндолимфа и купула сдвигались в
канале ампулофугально, активность
таких единиц тормозилась. При раздражении
вертикальных каналов возбуждающие
эффекты возникали при ускорениях,
вызывающих ампулофугальные токи, и
тормозные — при ускорениях, вызывающих
ампулопетальные токи эндолимфы. Эдриан
также описал единицы, которые не
обладали фоновой активностью, но
отвечали ритмическими разрядами
при действии ускорения.
Таким
образом было показано, что смещения
отолитов или отклонения купулы
приводят к изменениям частоты импульсации
вестибулярных нейронов, В последующих
работах более детальному анализу
была подвергнута активность нейронов,
отвечавших на раздражение горизонтальных
полукружных каналов (Eckel, 1954; Duensing,
Schaefer, 1958; Shimazu, Precht, 1965).
Шимазу
и Прехт (Shimazu, Precht, 1965), использовавшие
в качестве стимула ускорения,
обнаружили в верхнем и медиальном
вестибулярных ядрах кошки следующие
типы нейронов.
тип
реагировал увеличением частоты разрядов
на вращение в ипсилатеральную сторону
и снижением частоты на вращение в
контрлатеральную сторону (205 нейронов).
Этот тип нейронов обнаружен также в
вестибулярном нерве и ретикулярной
формации продолговатого мозга кошки
(Gernandt, 1949; Gernandt,
6*
83
Thulin,
1952), в вестибулярных ядрах кролика
(Eckel, 1954; Duensing, Schaefer, 1958).
тип
реагировал противоположно первому
типу. Частота импульсации снижалась
при вращении в ипсилатеральную и
увеличивалась при вращении в
контрлатеральную сторону (90 нейронов).
Единицы II типа в вестибулярном нерве
не обнаружены (Gernandt, 1949). Они описаны в
ретикулярной формации продолговатого
мозга кошки (Gernandt, Thulin, 1952) и вестибулярных
ядрах кролика (Duensing, Schaefer, 1958).
тип
нейронов отвечал учащением импульсации
на вращение в обе стороны (10 нейронов).
Найден в вестибулярном нерве кошки
(Gernandt, 1949) и вестибулярных ядрах кролика
(Eckel, 1954; Duensing, Schaefer, 1958).16
тип
характеризовался торможением активности
при вращении в обоих направлениях
(1 нейрон). Имеется в вестибулярном
нерве кошки и вестибулярных ядрах
кролика (Gernandt, 1949; Eckel, 1954; Duensing, Schaefer,
1958). Этот тип встречается редко. Гернандт
обнаружил 5 единиц из 100, Эккель — 1
единицу из 22.
Аналогичные
типы единиц были выявлены и при действии
адекватных раздражителей отолитов
(Егоров, 1967). Дюнзинг и Шефер (Duensing,
Schaefer, 1958) описали V тип нейронов, различно
реагирующих на вестибулярную
стимуляцию. Егоров (1967) считает, что
такие нейроны относятся к ретикулярной
формации.
В
I типе (Shimazu, Precht, 1965) выделена тоническая
группа нейронов, обладающая фоновой
ритмической активностью 2— 30 имп./сек.,
и кинетическая группа с очень редкойменее
(0.5имп./сек.) или полностью отсутствующей
фоновой импульсацией. Импульсная
активность у кинетических нейронов
появлялась только при действии ускорения.
Тонические нейроны имели более низкий
дорог (0.93 +0.55 град, /сек.2)
по сравнению с кинетическими(4.65 + + 1.84
град./сек,2).
Во время ускорения частота импульсации
тонических нейронов постепенно
нарастала до максимального уровня (для
данного ускорения) и по достижении
вращающимся стендом постоянной
угловой скорости медленно возвращалась
к исходному уровню.
Кинетические
нейроны в отличие от тонических
реагировали на ускорение резким
увеличением частоты импульсов. При
действии ускорений возрастающей
величины как тонические, так и кинетические
нейроны увеличивали частоту импульсации
по экспоненциальному закону (2/3
изученных единиц). Во время ускорения
тенденция к адаптации не отмечалась.
На стоп-стимул у большинства нейронов
частота импульсации снижалась, но у
части после торможения активности
имели место волнообразные изменения
частоты импульсации. Эти постстимульные
модуляции частоты
16 Описанные
три типа нейронов обнаружены также и
при адекватном раздражении передних
вертикальных каналов (Markham, 1968).
84
авторы
связывают с колебаниями купулы, допуская,
что купулЯ не всегда реагирует как
сверхкритически демпфированный маятник
и при некоторых условиях проявляет
качества демпфированного осциллятора.
Каким
образом тонические и кинетические
нейроны связаны с первичными вестибулярными
афферентами? Для решения этого вопроса
Прехт и Шимазу (Precht, Shimazu, 1965) наряду с
адекватным-раздражением полукружных
каналов применили методику отведения
фокальных потенциалов при электростимуляции
вестибулярного нерва. Фокальный
потенциал вестибулярных нейронов
состоит из нескольких компонентов: 1)
положительной или положительно-отрицательной
волны Р, являющейся ответом первичных
вестибулярных волокон; 2) второй
отрицательной волны N±
— моносинаптического
ответа вестибулярных нейронов; и 3)
следового отрицательного компонента
N2
—
полисинаптического ответа вестибулярных
нейронов.
У кинетических
нейронов сильнее выражен моносинаптический
компонент фокального ответа, а у
тонических — полисинаптиче- ский
компонент. О моносинаптической связи
кинетических нейронов с первичными
вестибулярными афферентами
свидетельствовали также короткая
синаптическая задержка (0.4—0.8 мсек.) и
небольшие колебания длительности
фокальных ответов (1.2— 2.0 мсек.). У
тонических нейронов, связанных с
первичными афферентами полисинаптически,
длительность ответов колебалась в
более широких пределах (1.5—8.6 мсек.),
что, возможно, обусловлено относительно
медленным развитием деполяризации
тонических нейронов по сравнению с
кинетическими.
Нейроны
I типа через комиссуральные волокна
получают тормозные влияния с
контрлатерального лабиринта. Эти
тормозные влияния полностью выпадают
после перерезки комиссуральных волокон.
Однако возбуждающие эффекты на более
сильную электростимуляцию VIII нерва,
осуществляемые через ретикулярную
формацию, сохраняются (Shimazu, Precht, 1966).
При раздражении контрлатерального
лабиринта средний скрытый период
тормозного эффекта тонических
нейронов I типа равняется 4.1 мсек., а
кинетических — 2.2 мсек. Это позволяет
сделать вывод о более прямых связях
контрлатерального лабиринта с
кинетическими нейронами I типа (Kasahara
et al., 1968). Изучение ТПСП показало, что
эти связи являются дисинаптическими.
Первичные вестибулярные афференты
моносинаптически возбуждают тормозные
комиссуральные нейроны медиального
ядра своей стороны, которые в свою
очередь моносинаптически вызывают
ТПСП в кинетических нейронах
медиального ядра противоположной
стороны (Mano et al., 1968). Нейроны, моносинаптически
связанные с первичными вестибулярными
волокнами, локализуются в основном в
ростральной части медиального ядра
(Wilson et al., 1968b).
85
В
отличие от нейронов I типа нейроны II
типа на электрораздражение
контрлатерального VIII нерва отвечают
возбуждением. Перерезка комиссуральных
волокон снимает этот эффект. Шимазу и
Прехт (Shimazu, Precht, 1966) предлагают следующую
схему взаимодействия нейронов
вестибулярных ядер. Возбуждение
рецепторов горизонтального
полукружного канала вызывает активацию
вестибулярных нейронов I типа в ядрах
своей стороны.17
Часть из этих нейронов, являющихся
вставочными, отдает комиссуральные
волокна к нейронам II типа ядер
противоположной стороны, которые
оказывают тормозное действие на нейроны
I типа своей стороны. G помощью такого
механизма усиливается различие в
уровнях возбуждения вестибулярных
ядер обеих сторон. Описанная схема
взаимодействия полностью соответствует
схеме Г. И. Гор- гиладзе (19646, 1966а, 19666),
который предложил ее, исходя из
экспериментов с поляризацией лабиринтов
постоянным током.
ВЕСТИБУЛЯРНЫЙ
КОНТРОЛЬ СПИНАЛЬНЫХ РЕФЛЕКСОВ
Вестибуло-спинальные
влияния осуществляются в основном
через вестибуло-спинальный путь, берущий
начало в латеральном ядре, через
вестибуло-спинальные связи медиального
ядра, через ретикуло-спинальные волокна,
а также нисходящий медиальный продольный
пучок (Brodal, 1967b; Precht et al., 1967; Gernandt, 1968b;
Wilson et al., 1968a). Скорость проведения
возбуждения по волокнам
вестибуло-спинального тракта колеблется
от 24 до 140 м/сек. (Ito et al., 1964). Вестибулярный
аппарат управляет спинальными
двигательными реакциями путем посылки
импульсов к а- и у-мотонейронам (см.
обзор: Brodal et al., 1962; Gernandt, 1967), а также к
вставочным нейронам (интернейронам)
спинного мозга (Erulkar et al., 1966). В ПСП
а-мотонейронов, вызванные раздражением
лабиринта, имеют как моно-, так и полиси-
наптический механизм возникновения.
Продолжительность латентного периода
ТПСП (6—7 мсек.) указывает на их полисинапти-
ческую природу (Шаповалов и др., 1966).
На
уровне вставочных нейронов спинного
мозга обнаружено сложное взаимодействие
раздражителей различных модальностей,
влияющих на позу и равновесие животного.
В шейном и поясничном отделах спинного
мозга кошки обнаружены три популяции
вставочных нейронов: 1) отвечающих
только на вестибулярную стимуляцию;
2) реагирующих на совместное раздражение
вестибулярного нерва и дорсального
корешка и 3) реагирующих на совместное
раздражение вестибулярного нерва и
моторной коры.
17 Нейроны
типа I перекрестно влияют на мотонейроны
и прямо — ша вставочные нейроны ядра
отводящего нерва (Precht et al*, 1969).
86
Некоторые
нейроны отвечают на раздражители всех
модальностей. Гистологически показано,
что вставочные нейроны, возбуждающиеся
раздражением вестибулярного нерва,
локализуются в медиальных частях
серого вещества спинного мозга, в
области окончания аксонов вестибуло-
и ретикуло-спинального трактов. Нейроны,
отвечающие на совместную стимуляцию
вестибулярного нерва и дорсального
корешка, локализованы в центре серого
вещества спинного мозга, а нейроны,
реагирующие на совместное раздражение
вестибулярного нерва и моторной коры,
— в латеральных его частях, т. е. в
конечных областях соответствующих
трактов. Стимуляция вестибулярного
нерва сопровождается ответами как
вентральных (моторных), так и дорсальных
(чувствительных) корешков спинного
мозга. Следовательно, часть вставочных
нейронов, на которые проецируется
вестибулярная система,’может
пресинаптически влиять на афферентный
спинномозговой вход (Erulkar et al., 1966).
Наряду
с вестибулярным контролем спинальных
рефлексов существуют спинальные влияния
на вестибулярные ядра, изменяющие
функциональное состояние и активность
нейронов (Вго- dal et al., 1962). Например,
электростимуляция нервов передних и
задних конечностей, а также кожных
нервов оказывает облегчающие и
тормозные эффекты на спонтанноактивные
нейроны латерального ядра Дейтерса
(Wilson et al., 1966, 1967),
ВЕСТИБУЛО-МОЗЖЕЧКОВЫЕ
ФУНКЦИОНАЛЬНЫЕ
ОТНОШЕНИЯ
Мозжечок
получает как первичные, так и вторичные
вестибулярные волокна, а также посылает
аксоны к вестибулярным ядрам
продолговатого мозга, которые отходят
от клеток коры червя, фастигиального
ядра и особенно обильно снабжают
латеральное вестибулярное ядро
Дейтерса (Brodal et al., 1962; Вго- dal, 1964, 1967b).
Проекционные зоны вестибулярного
нерва, определенные методикой
вызванных потенциалов, локализованы
в nucleus fastigii, uvula, lingula и флоккулонодулярной
доле мозжечка" (Dow, Moruzzi, 1958).
Микроэлектродными
исследованиями показано, что импульсы
вестибулярного аппарата изменяют
активность клеток Пуркинье (Чжан
Сян-дун, Костюк, 1960; Llinas et al., 1967), которые
в свою очередь контролируют активность
вестибулярных нейронов (Ito et al., 1968).
Нейроны коры и ядер мозжечка обрабатывают
информацию об ускорениях. Корреляционный
анализ изменений межспайковых
интервалов позволил сделать заключение,
что в мозжечке имеются нейроны,
выполняющие операцию слежения за
изменениями ускорения (Вентцель и
др., 1969).
Мозжечок
контролирует различные вестибулярные
рефлексы (нистагм глаз, спинальные
рефлексы и т. п.), которые изменяются
а?