

Лысак Микробиология
.pdfвращается в линейную. Это было неожиданным, так как для актиномицетов уже существовали физические кольцевые карты. Однако выяснилось, что хромосомы актиномицетов оказались «псевдокольцевыми»: они были замкнуты не за счет непрерывного перехода цепочки ДНК правого полукружия хромосомы в левое, а за счет взаимодействия белковых молекул, расположенных на свободных концах ДНК линейной хромосомы, и замыкающих их. Протеазы разрывают эту связь. Таким образом, у актиномицетов другой тип организации хромосомы, чем у боррелий. Линейная хромосома была найдена ещё и у фитопатогенных бактерий вида Rhodococcus fascians.
Отметим основные моменты репликации кольцевой бактериальной хромосомы, поскольку этот тип организации является основным для хромосом бактерий.
Теоретически возможны три механизма репликации (рис. 23):
1.Консервативный механизм, при котором сохраняется целостность всей родительской двойной спирали (не происходит раскручивания спирали), и она является матрицей для синтеза себе подобной. Дочерняя двойная спираль полностью образуется из нового материала, а родительская как таковая сохраняется.
2.Дисперсивный механизм, в соответствии с которым родительская молекула ДНК распадается на фрагменты, а синтез новых цепей происходит на фрагментах, которые затем крест-накрест объединяются с отрезками нового материала. Каждая полинуклеотидная цепь в этом случае должна была бы состоять из чередующихся отрезков старого и нового материала.
3.Полуконсервативный механизм предполагает, что родительская двойная спираль раскручивается, и каждая полинуклеотидная цепь служит матрицей для синтеза новой комплементарной цепи. Таким образом, новая двойная молекула оказывается «гибридом» старой
ивновь синтезированной цепи.
консервативный дисперсивный полуконсервативный
Рис. 23. Механизмы репликации ДНК
70

В 1957 году М.Меселсон и Ф.Сталь экспериментально доказали, что репликация хромосомной ДНК у бактерий происходит по полуконсервативному механизму. Для доказательства этого бактерии E.coli выращивали на среде в присутствии хлористого аммония, содержащего тяжелый изотоп азота (15N). Через некоторое время бактерии переносили на среду с легким азотом (14N). До и после пересева через определенные промежутки времени отбирали пробы бактерий, лизировали их, из клеток выделяли ДНК и определяли ее плотность методом равновесного центрифугирования в градиенте плотности хлористого цезия. Параллельно бактерии выращивали в присутствии только легких или только тяжелых изотопов азота и проводили те же процедуры, что и с опытными культурами E.coli.
Оказалось, что препараты ДНК, выделенные из бактерий, выращенных на средах с разными формами азота, отличаются по плотности. ДНК, выделенная из бактерий, выращенных на среде с 15N, более тяжелая, чем ДНК бактерий, выращенных на среде с 14N. ДНК, выделенная же из бактерий, первоначально растущих, на среде с 15N, а затем перенесенных на среду с 14N, имела промежуточную плотность, т.е. была «полутяжелой» или «гибридной» ДНК. Общее количество такой полутяжелой ДНК оставалось постоянным на протяжении нескольких поколений (генераций), тогда как количество «легкой» ДНК увеличилось. Меселсон М. и Сталь Ф. сделали вывод, что в молекуле «полутяжелой» ДНК в составе пуриновых и перимидиновых оснований одна цепь содержит 15N, а вторая – 14N (рис. 24).
легкая 14N-ДНК гибридная ДНК
тяжелая 15N-ДНК легкая 14N-ДНК гибридная ДНК после трех генераций (после одной генерации)
Рис. 24. Схема опыта М.Меселсона и Ф.Сталя
Результаты этих экспериментов не объясняются ни с консервативным, ни с дисперсивным механизмом репликации ДНК, т.к. при консервативном механизме выявились бы только полосы тяжелой и легкой ДНК, а при дисперсивном – только полосы гибридной ДНК (и после нескольких генераций). Для доказательства того, что «полутяжелая» ДНК состоит из одной нити содержащей 15N и одной нити содержащей 14N они ее подвергали «плавлению» (нагревали до 100ºС и быстро охлаждали). При этом образовывались одноцепочечные ДНК
71
(разрывались водородные связи). Полученный препарат центрифугировали в градиенте плотности хлористого цезия, после чего были выявлены две полосы: типичная для одноцепочечной ДНК с 14N и типичная для такой же ДНК с 15N.
Таким образом, М.Меселсон и Ф.Сталь доказали, что репликация хромосомной ДНК у бактерий осуществляется по полуконсервативному механизму.
Молекулярные механизмы репликации бактериальной ДНК сходны в общих чертах с таковыми у других организмов. Для того, чтобы произошла репликация ДНК по полуконсервативному механизму необходимо, чтобы двухцепочечная молекула ДНК расплелась с образованием одноцепочечных фрагментов. В этом процессе участвует несколько ферментов, основными из них являются:
1)ДНК-геликазы, перемещающиеся по цепи ДНК в направлении 5/ → 3/, и перемещающиеся в направлении 3/ → 5/;
2)SSB-белки (single strand binding – белки, связывающиеся с однонитевой ДНК), которые связываются с однонитевой ДНК и препятствуют ее ренатурации;
3)ДНК-гиразы, или топоизомеразы белки, которые снимают напряжение при раскручивании кольцевых молекул ДНК и способствуют ее расплетанию.
На образовавшихся однонитевых участках ДНК идет синтез комплементарных цепей. В этом процессе участвуют ферменты ДНКполимеразы. У бактерий E.coli синтезируются три типа ДНКполимераз (I, II и III). Главную роль в репликации хромосомной ДНК
убактерий E.coli играет ДНК-полимераза III.
Доказано, что синтез ДНК протекает только в направлении от 5/ к 3/-ОН концу (рис. 25). Однако, цепи ДНК противоположно направлены и поэтому синтез одной из дочерних цепей осуществляется непрерывно с помощью ДНК-полимеразы III в направлении 5/ → 3/. ДНКполимераза III перемещается по цепи 3/ → 5/, в направлении раскрывания репликативной вилки и синтезирует новую цепь. Эта цепь на-
зывается ведущей или лидирующей.
Копия второй цепи (5/ → 3/) называется запаздывающей и она синтезируется из фрагментов ДНК размером 1000 – 2000 нуклеотидов. Они называются фрагментами Оказаки. Для синтеза фрагментов Оказаки необходима “затравка” или праймер. В качестве затравки выступает короткая цепь РНК, которая синтезируется с помощью ДНК-зависимой РНК-полимеразы на матрице ДНК. К этой затравке ДНК-полимераза III присоединяет дезоксирибонуклеотиды и образуются фрагменты Оказаки. РНК-праймеры удаляются за счет активно-
72

сти ДНК-полимеразы I, после чего фермент лигаза сшивает отдельные фрагменты Оказаки друг с другом и восстанавливает целостность новой цепи.
ДНК-полимераза
на лидирующей цепи
лидирующая цепь
родительская двухцепочечная ДНК
РНК-затравка
ДНК-геликаза
фрагменты Оказаки
ДНК-праймаза
SSB-белок
запаздывающая цепь
ДНК-полимераза на запаздывающей цепи
Рис. 25. Репликации ДНК у бактерий E.сoli в соответствии с полуконсервативным механизмом
Репликация всего кольца ДНК может происходить как в одном, так и в двух противоположных направлениях двойной спирали. Такие механизмы репликации предполагают наличие одной или двух репликативных вилок на одной молекуле ДНК.
В качестве примера можно указать, что время удвоения хромосомы бактерий E.coli занимает приблизительно 40 минут. Однако в благоприятных условиях деление клеток происходит за 20 минут. Это значит, что новый цикл репликации дочерних хромосом начинается еще до того, как заканчивается предыдущий.
Процесс репликации тесно связан с ростом и делением бактериальной клетки. Обычно деление бактериальной клетки по времени осуществляется после завершения цикла репликации молекулы ДНК. Однако, в интенсивно растущих культурах репликация ДНК может опережать процесс деления клетки и нередко в клетке содержится ДНК в 4 – 8 раз больше чем масса одной хромосомы. Уже отмечалось, что между бактериальной ДНК и цитоплазматической мембраной существует физическая связь, так как бактериальную ДНК можно обнаружить в мембранных фракциях после центрифугирования лизата, а так же с помощью электронной микроскопии удалось визуализиро-
73
вать места прикрепления хромосомы к впячиваниям мембраны (мезосомам).
Связь бактериальной хромосомы или плазмиды с цитоплазматической мембраной, играет существенную роль в регуляции их репликации. Существуют две модели, объясняющие регуляцию репликации бактериальной ДНК. Согласно модели, предложенной Ф.Жакобом, С.Бреннером и Ф.Кьюзеном (1963), структура, способная самостоятельно реплицироваться, называется репликоном, что относится к хромосомам и плазмидам бактерий. Репликон должен иметь кольцевую форму и реплицироваться не по частям, а как единое целое. Согласно этой модели репликон должен быть прикреплен к цитоплазматической мембране и обязательно обладать двумя специфическими детерминантами или генами – структурным геном и геном-реплика- тором (или оператором репликации). При росте клетки от мембраны поступает сигнал на структурный ген и активирует его. Происходит синтез специфического белка-инициатора, который действует на генрепликатор, что приводит к началу процесса репликации, который продолжается вдоль всего репликона и заканчивается копированием всей его структуры. После репликации ДНК поступает обратный сигнал на мембрану, инициируя деление клетки. Данная модель получила название модели позитивной регуляции репликации.
Кроме этой модели существует модель негативной регуляции репликации (Р.Притчард, П.Барт, Дж.Коллинз, 1969). В соответствии с этой моделью в составе репликона есть ген, отвечающий за синтез белка-репрессора, который при высокой концентрации негативно действует на инициацию репликации, в малой концентрации не влияет на этот процесс. По мере роста клетки, концентрация репрессора снижается и создается возможность репликации хромосомы или плазмиды.
74
Глава 3. ВИРУСЫ: ОБЩАЯ ХАРАКТЕРИСТИКА
Вирусы – мельчайшие объекты жизни, имеющие неклеточное строение и не способные к проявлению каких-либо признаков живого вне живых клеток. Первым открытым вирусом был вирус мозаичной болезни табака, обнаруженный русским ученым Д.И.Ивановским в
1892 году.
Каждый вирус в своем онтогенезе проходит 2 стадии:
-внеклеточную, когда вирус находится в состоянии покоя, и получившую название вирион. В таком состоянии он находится в условиях окружающей среды;
-внутриклеточную, в течение которой происходит весь цикл репродукции в клетках хозяина.
Каждый вирион представлен двумя основными компонентами – нуклеиновой кислотой и белком, что позволяет называть вирусы неклеточными формами жизни. Отличия вирусов от клеточных организмов представлены в табл.4.
|
|
|
|
Таблица 4 |
|
|
Отличие вирусов от клеточных организмов |
||||
|
|
|
|
|
|
Свойства |
|
Вирусы |
Прокариоты |
Эукариоты |
|
|
|
|
|
|
|
Клеточная орга- |
|
- |
+ |
+ |
|
низация |
|
|
|||
|
|
|
|
|
|
Тип нуклеиновой |
|
ДНК или РНК |
ДНК + РНК |
ДНК + РНК |
|
кислоты |
|
|
|
|
|
Автономный ме- |
|
- |
+ (кроме некото- |
+ |
|
таболизм |
|
|
рых риккетсий) |
|
|
|
|
|
|
||
Рост на питатель- |
|
- |
+ (кроме риккет- |
+ |
|
ных средах |
|
сий) |
|
||
|
|
|
|
||
Бинарное деление |
|
- |
+ |
+ |
|
|
|
|
|
|
|
3.1. Строение и химический состав вирусных частиц
Уже было отмечено, что вирусная частица состоит из нуклеиновой кислоты (ДНК или РНК) и белка. Белковая оболочка, которая окружает нуклеиновую кислоту, называется капсидом. Капсид каждого вируса состоит из отдельных субъединиц – капсомеров. Капсиды некоторых вирионов окружены дополнительной мембраной. Если вирус имеет мембрану, то говорят, что он «в оболочке» или окружен суперкапсидом, при отсутствии мембраны вирус называют «раздетым» или «голым».
75
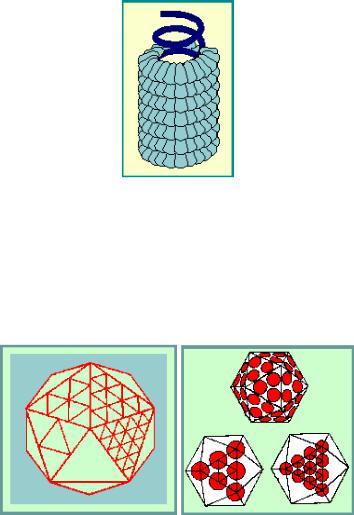
Капсид чаще всего имеет симметричное строение. Различают два типа симметрии – спиральную и кубическую. При спиральной симметрии капсида вирусная нуклеиновая кислота образует спиральную (или винтообразную) фигуру, полую внутри, и субъединицы белка (капсомеры) укладываются вокруг нее тоже по спирали (трубчатый капсид) (рис. 26). Примером вируса со спиральной симметрией капсида является вирус табачной мозаики, который имеет палочковидную форму, а его длина составляет 300 нм с диаметром 15 нм. В состав вирусной частицы входит одна молекула РНК размером около 6000 нуклеотидов. Капсид состоит из 2000 идентичных субъединиц белка, уложенных по спирали.
Рис. 26. Строение вируса со спиральной симметрией
При кубической симметрии вирусная нуклеиновая кислота уложена плотно (свернута в клубок), а белковые молекулы окружают ее, образуя многогранник (икосаэдр). Икосаэдр – многогранник с двадцатью треугольными гранями, имеющий кубическую симметрию и приблизительно сферическую форму (рис. 27). К икосаэдрическим вирусам относятся вирус простого герпеса, реовирусы и др.
Рис. 27. Строение вируса с кубической симметрией
В зависимости от типа симметрии вирусы подразделяют на: вирусы со спиральным типом симметрии, вирусы с кубическим типом
76
симметрии и сложные вирусы, имеющие оба типа симметрии и состоящие из икосаэдрической головки и хвоста. Примером сложных вирусов являются колифаги Т2, Т4 (т.е. бактериофаги, инфицирующие клетки бактерий E.coli) и др. У некоторых сложных вирусов икосаэдрический капсид заключает в себе трубчатый нуклеокапсид.
Взависимости от того, какой тип нуклеиновой кислоты содержится в вирусной частице, их подразделяют на 2 группы:
- ДНК-содержащие, имеющие в качестве генетического материала либо одно-, либо двухцепочечную ДНК, которая может быть как линейной, так и кольцевой. Примером ДНК-содержащих вирусов являются колифаги Т2, Т4, λ; вирус простого герпеса; вирус оспы и др.
- РНК-содержащие вирусы, генетическая информация которых закодирована в РНК. РНК также может быть как одно-, так и двухцепочечной. Вирусы с одноцепочечной РНК можно разделить, в свою очередь, на 2 типа: с «плюс»-цепью РНК и с «минус»-цепью РНК. У вирусов первого типа цепь РНК может функционировать в клеткехозяине непосредственно как информационная РНК, тогда как у вирусов второго типа на «минус»-цепи должна сначала с помощью клеточных РНК-полимераз синтезироваться «плюс»-цепь РНК, которая и служит информационной РНК. Примером РНК-содержащих вирусов являются реовирусы, вирус гриппа, ретровирусы и др.
Взависимости от организма хозяина выделяют вирусы, поражающие животных и человека, растения и микроорганизмы.
Вирусы растений иначе называются фитопатогенными вирусами. Эти вирусы попадают внутрь растительных клеток через повреждения или с помощью переносчиков насекомых или нематод. Фитопатогенные вирусы вызывают у растений множество болезней. Особенно большой вред приносят вирусы, поражающие картофель.
Учеловека вирусы вызывают множество заболеваний, включая оспу, грипп, корь, свинку, инфекционный гепатит, желтую лихорадку, полимиелит, герпес, бешенство, СПИД, раковые заболевания и др. Многие вирусные заболевания у человека и животных можно предупредить путем иммунизации. Вирусные заболевания поддаются лечению с трудом, так как вирусы не чувствительны к большинству антибиотиков. Вирусы животных и человека передаются при контакте, либо с помощью насекомых-переносчиков, а также через объекты окружающей среды.
Вирусы бактерий называются бактериофагами. Вероятно в природе не существуют бактерии, которые не поражались бы одним из типов
77
бактериофагов. Бактериофаги могут наносить вред производству, основанному на жизнедеятельности бактерий.
3.2. Строение бактериофагов. Взаимодействие бактериофагов с чувствительными клетками бактерий
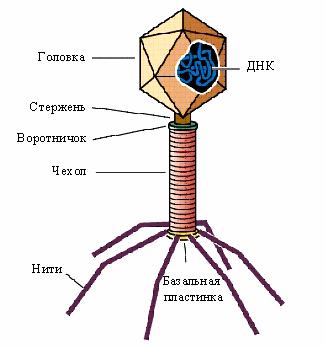
Строение бактериофагов можно рассмотреть на примере колифага Т4, электронная микрофотография которого была получена одной из первых. Этот бактериофаг, как и все Т-четные колифаги относится к сложным вирусам, т.е. он состоит из икосаэдрической головки диаметром 650 Å и длиной 950 Å, и отростка или хвоста (рис. 28). Капсид головки состоит из капсомеров, внутри него находится плотно упакованная двухцепочечная линейная ДНК и фермент транскриптаза в неактивном состоянии. Отросток фага имеет сложное строение. Он состоит из полого стержня, покрытого чехлом, который заканчивается базальной пластинкой с шипами и нитями. Все структуры отростка состоят из белков. В области базальной пластинки находится фермент бактериофаговый лизоцим, который способен разрушать пептидогликан муреин клеточной стенки бактерий. Здесь же имеется АТФ-аза, которая регенерирует энергию для сокращения чехла отростка бактериофага.
Рис. 28. Строение бактериофага Т4
78
Другие из изученных бактериофагов имеют более простое строение. В зависимости от формы зрелых фаговых частиц различают следующие морфологические типы бактериофагов:
-состоящие из икосаэдрической головки и спирального хвоста с сократимым чехлом (Т-четные колифаги);
-состоящие из икосаэдрической головки и длинного гибкого несократимого отростка (колифаги Т1 и Т5);
-нитчатые бактериофаги (колифаг fd);
-состоящие из икосаэдрической головки с коротким несократимым отростком (колифаги Т3 и Т7, фаг Р22 бактерий Salmonella typhimurium);
-состоящие из икосаэдрической головки без отростка. Большинство бактериофагов содержит двухцепочечную ДНК, но
охарактеризованы и бактериофаги с одноцепочечной ДНК (колифаг М13) и с одноцепочечной РНК (колифаги fd, Qβ, R17).
В зависимости от способа размножения в чувствительной клетке бактериофаги подразделяются на 2 группы: вирулентные и умеренные.
Вирулентные фаги всегда лизируют зараженные ими бактерии и имеют только один путь развития – литический цикл. Умеренные фаги могут вести себя по-разному: после проникновения в клетку нуклеиновая кислота фага либо вовлекается в литический цикл, либо вступает с клеткой-хозяином в своего рода симбиотические отношения, т.е. встраивается в хромосому бактериальной клетки (превращается в профаг) и передается всему потомству данной клетки. Бактерии, которые содержат профаг, называются лизогенными.
Рассмотрим взаимодействие вирулентных фагов с чувствитель-
ной клеткой хозяином на примере колифага Т4.
При смешивании взвеси фаговых частиц с суспензией бактерий фаговые частицы в результате случайных столкновений с клетками бактерий прикрепляются к последним (адсорбируются). Адсорбция происходит на рецепторах, имеющихся в наружной мембране бактерий E.coli. За адсорбцией следует стадия инъекции или введения ДНК в клетку. Бактериофаговый лизоцим разрушает клеточную стенку бактерий и с помощью энергии, регенерируемой АТФ-азой, происходит сокращение чехла бактериофага. При этом прокалывается цитоплазматическая мембрана, полый стержень входит в бактериальную клетку и ДНК фага впрыскивается в нее.
79