

Адреналін гальмує синтез глікогену
Адреналін не тільки стимулює розпад глікогену, але й одночасно гальмує його синтез в печінці із глюкози, що сприяє максимальному потраплянню глюкози в кров. Як видно на Рис. 2.1, зв’язування адреналіну на поверхні клітин печінки та наступне утворення сАМР стимулюють каталізуючий протеїнкіназою процес фосфорилювання глікоген-синтази, в результаті якого активна дефосфорильована форма глікоген-синтази перетворюється в неактивну фосфорильовану форму. Таким чином, ланцюг реакцій, який призводить до зменшення глікоген-синтазної активності, має той же самий механізм, що і розпад глікогену з утворенням вільної глюкози в крові.
Внаслідок увесь доступний глікоген, глюкозо-6-фосфат та інші попередники йдуть на утворення вільної глюкози, яка надходить у кров; цим досягається максимальне забезпечення м'язів паливом і тим самим здійснюється підготовка організму до критичної ситуації. Адреналін діє не тільки на печінку, але і на скелетні м'язи та серце, де він сприяє розпаду глікогену також шляхом стимуляції м'язової фосфорилази через утворення сАМР.
Р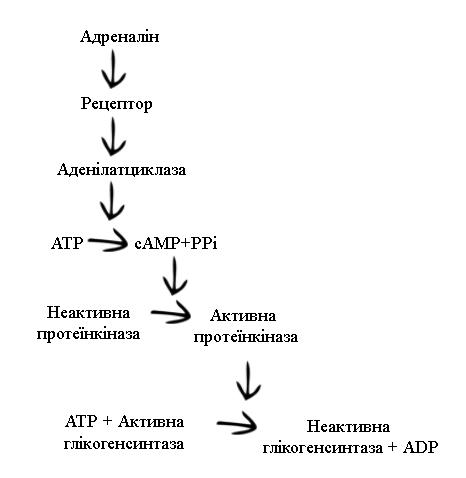 ис.
2.1
Гальмування
синтезу глікогену адреналіном.
Протеїнкіназа, активована циклічним
АМР, каталізує фосфорилювання
глікоген-синтази, переводячи таким
чином фермент в неактивну форму.
ис.
2.1
Гальмування
синтезу глікогену адреналіном.
Протеїнкіназа, активована циклічним
АМР, каталізує фосфорилювання
глікоген-синтази, переводячи таким
чином фермент в неактивну форму.
Оскільки в м'язах і серці відсутня глюкозо-6-фосфатаза, продуктом розщеплення глікогену в цих органах є не глюкоза крові, а молочна кислота, яка утворюється з глюкозо-6-фосфату під часі гліколізу. Таким чином, стимуляція розпаду глікогену в м'язах веде до збільшення швидкості гліколізу та утворенню АТР, що забезпечує швидке зростання м'язової активності.
Фосфодіестераза інактивує циклічний аденозинмонофосфат
Д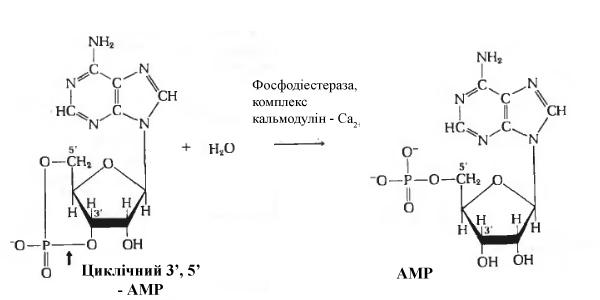 о
тих пір, доки тварина відчуває себе в
небезпеці і мозковий шар його наднирників
секретує в кров адреналін, аденілатциклазна
система печінки залишається повністю
активованою. В результаті концентрація
сАМР в клітинах-мішенях підтримується
на відносно високому рівні, що забезпечує
високу швидкість розпаду глікогену.
Але як тільки небезпека зникає, секреція
адреналіну припиняється і його вміст
в крові швидко падає в результаті
ферментативного розщеплення в печінці.
По мірі того, як рецептори адреналіну
стають незайнятими, аденілатциклаза
повертається в неактивний стан та
утворення сАМР припиняється. сАМР який
залишився руйнується під дією
фосфодіестерази (Рис. 3.1) – ферменту,
який каталізує гідроліз 3’-фосфатного
зв’язку в сАМР з утворенням вільного
5’-аденозинмонофосфату (5’-АМР):
о
тих пір, доки тварина відчуває себе в
небезпеці і мозковий шар його наднирників
секретує в кров адреналін, аденілатциклазна
система печінки залишається повністю
активованою. В результаті концентрація
сАМР в клітинах-мішенях підтримується
на відносно високому рівні, що забезпечує
високу швидкість розпаду глікогену.
Але як тільки небезпека зникає, секреція
адреналіну припиняється і його вміст
в крові швидко падає в результаті
ферментативного розщеплення в печінці.
По мірі того, як рецептори адреналіну
стають незайнятими, аденілатциклаза
повертається в неактивний стан та
утворення сАМР припиняється. сАМР який
залишився руйнується під дією
фосфодіестерази (Рис. 3.1) – ферменту,
який каталізує гідроліз 3’-фосфатного
зв’язку в сАМР з утворенням вільного
5’-аденозинмонофосфату (5’-АМР):
Рис. 3.1 Дія фосфодіестерази на сАМР. Фосфодіестераза багатьох тканин активується іонами Са2+. Цей ефект непрямий; спочатку іони Са2+пов’язуються з регуляторним білком кальмодуліном, а потім комплекс Са2+-кальмодулін приєднуються до фосфодіестерази, викликаючи тим самим активацію ферменту.
По мірі зменшення складу сАМР в цитозолі відбувається вивільнення сАМР, який пов’язаний з регуляторними субодиницями протеїнкінази. В результаті регуляторні субодиниці поєднуються знову з каталітичними в результаті чого протеїнкіназа переходять в неактивну форму. Фосфорильована форма кінази фосфорилази в подальшому підлягає дефосфорилюванню так як і фосфорилаза а, під дією фосфатази фосфорилази. Все це повертає систему глікогенолізу в вихідний стан. Одночасно відбувається ре активація глікоген-синтази шляхом її дефосфорилювання. Характерною що вона інгібується кофеїном та теофіліном – алкалоїдами, які в великих кількостях знаходяться в великих кількостях в каві та чаї. Ці алкалоїди пролонгують або посилюють дію адреналіну шляхом зниження швидкості розпаду сАМР. В цілому ряді тканин фосфодиестераза активується іонами Са2++. Цей ефект пояснюється зв’язуванням іонів Са2++ зі специфічними Са2++-зв’язуючим білком, який називається кальмодуліном. Комплекс Са2++-кальмодулін приєднується до фосфодиестерази та активує її. Кальмодулін представляє собою нещодавно відкритий білок, який широко розповсюджений в усьому тваринному світі. Майже у усіх тварин кальмодулін має одну й ту ж саму послідовність амінокислот. Концентрація іонів Са2++ в цитозолі регулює багато функцій клітин; в силу цього іони Са2+, подібно сАМР, відіграють важливу регуляторну роль як вторинний посередник. Кальмодулін бере учать в передачі сигналу, який викликає збільшення іонів Са2+ в цитозолі, шляхом утворення комплексу Са2+-регулючим білком, стимулює його активність.