

Механизмы мышечного сокращения
Структура миофибрилл и ее изменения при сокращении. Миофибриллы представляют собой сократительный аппарат мышечного волокна. В поперечнополосатых мышечных волокнах миофибриллы разделены на правильно чередующиеся участки (диски), обладающие разными оптическими' свойствами. Одни участки анизотропны, т. е. обладают двойным лучепреломлением.В обыкновенном свете они выглядят темными, а в поляризованном — прозрачными в продольном направлении и непрозрачными в поперечном. Другие участки в обыкновенном свете выглядят светлыми — они изотропны, т. е. не обладают двойным лучепреломлением (рис. 34, а). Анизотропные участки обозначают буквой Л, изотропные — буквой I. В середине диска Л различается светлая полоска Н, посередине диска I — темная полоска Z, представляющая собой тонкую мембрану, сквозь поры которой проходят миофибриллы. Благодаря наличию этой опорной структуры параллельно расположенные однозначные диски отдельных фибрнлл внутри одного волокна во время сокращения не смещаются по отношению друг к другу.
Современные представления о структуре миофибриллярпого аппарата основываются на исследованиях структуры мышечного волокна при помощи электронной микроскопии, рентгеноструктурного анализа, фазово-контрастной и интерференционной микроскопии в сочетании с гистохимическими методами.
Установлено, что каждая миофибрилла мышечного волокна диаметром около 1 мкм состоит в среднем из 2500 протофибрилл, представляющих собой удлиненные полимери-зованные молекулы белков миозина и актина. Миозиновые протофибриллы, или, как их принято обозначать, нити, вдвое толще актиповых. Их диаметр примерно 10 нм. В состоянии покоя мышечного волокна нити расположены в миофибрилле таким образом, что тонкие длинные актиновые нити входят своими концами в промежутки между толстыми и более короткими миозиновыми нитями (рис. 34, б). Благодаря этому диски / состоят только из актиновых нитей, а диски Л — из нитей миозина, а, возможно, еще и другого белка.
Светлая полоска Н представляет собой узкую зону, свободную от актиновых нитей. Мембрана Z, проходя через середину диска I, скрепляет между собой эти нити:
Важным компонентом ультрамикроскопической структуры миофибрилл являются также многочисленные поперечные мостики, соединяющие между собой миозиновые и актиновые нити. При сокращении мышечного волокна указанные нити не укорачиваются, а начинают «скользить» друг по другу: актиновые нити вдвигаются в промежутки между миозиновыми, в результате чего диски I укорачиваются, а диски Л сохраняют свой размер. Почти исчезает светлая полоска Н, так как актиновые нити при сокращении сближаются друг с другом своими концами (рис. 34, в). Причиной «скольжения» является химическое взаимодействие между актином и миозином в присутствий-ионов Са24 и АТФ. Наблюдается своего рода химическое «зубчатое колесо», как бы протягивающее одну группу нитей по другой. Роль «зубчиков» в этом процессе приписывают поперечным мостикам, обеспечивающим взаимодействие активных центров белков миозиновых и актиновых нитей.
Роль потенциала действия в возникновении мышечного сокращения. В естественных условиях деятельности скелетной мышцы инициатором ее сокращения является потенциал действия, распространяющийся при возбуждении вдоль поверхностной мембраны мышечного волокна.
Если кончик микроэлектрода при помощи микроманипулятора приложить к поверхности мышечного волокна лягушки в области диска I, то при нанесении очень слабого электрического стимула, вызывающего деполяризацию, диски I по обе стороны от мембраны Z начнут укорачиваться. При этом, однако, сокращение распространяется не в стороны, а в глубь волокна, вдоль диска I. Приложение слабого стимула к другим участкам миофибриллы подобного эффекта не вызывает. Из этого следует, что деполяризация поверхностной мембраны мышечного волокна в области дисков I является пусковым механизмом сократительного процесса.
Важным промежуточным звеном между деполяризацией мембраны и началом мышечного сокращения является проникновение в область миофибрилл свободных ионов Са2+. В состоянии покоя основная часть ионов Са2+ в скелетном мышечном волокне хранится в так называемом саркоплазматическом ретикулуме. Он представляет собой замкнутую систему внутриклеточных трубочек и цистерн, окружающих каждую миофибриллу.
В мембране саркоплазматического ретикулума локализованы две важнейшие транспортные системы, обеспечивающие накопление в ретикулуме ионов Ca2+ («секвестрация» — захват их из миоплазмы) и освобождение Са2+ из ретикулума при возбуждении.
Функцию кальциевого насоса выполняет так называемая Са - зависимая АТФ-аза (Са — АТФ-аза). Энергия, выделяющаяся при расщеплении АТФ, используется для секвестрации ионов Са2+ в ретикулум. Благодаря этому в покоящемся волокне концентрация свободных ионов Са2+ в цитоплазме поддерживается на очень низком уровне. Поступая внутрь ретикулума (главным образом в его продольные трубочки), ионы Са2+ частично связываются белковыми молекулами, устилающими внутреннюю поверхность его трубочек и цистерн. Концентрация свободных Са2+ в полости ретикулума близка к концентрации их в наружной среде, т. е. во внеклеточной жидкости.
В механизме освобождения ионов Ca2+ из ретикулума при возбуждении особую роль играет система поперечных трубочек (Т-система), представляющих собой впячивания поверхностной мембраны. Диаметр каждой трубочки около 0,05 мкм. На рис. 35 приведена схема продольного среза через быстрое мышечное волокно лягушки. Видно, что по обе стороны от поперечной трубочки расположены боковые (терминальные) цистерны ретикулума. Вместе с трубочкой они образуют так называемые триады. Мембрана поперечных трубочек по своим свойствам сходна с поверхностной мембраной; она содержит электровозбудимые натриевые каналы и способна к генерации и проведению потенциала действия. Во время возбуж
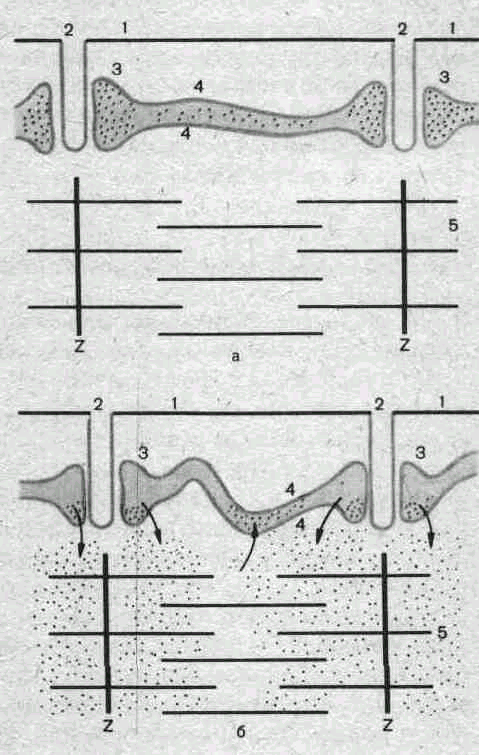
Рис. 35. Схематическое изображение взаимоотношений поверхностной мембраны (1),поперечных трубочек (2),боковых цистерн (3) и продольных трубочек (4) саркоплазматического ретикулума и миофибрилл (5) мышечного волокна.
а — в состоянии покоя, б — во время сокращения. Деполяризация мембраны и поперечных трубочек вызвала освобождение ионов Са2+ из боковых цистерн. Освободившийся Са2+ диффундирует по направлению к миофибриллам и частично захватывается продольными трубочками ретикулума.
Падения потенциал действия с поверхностной мембраны распространяется вдоль мембраны поперечных трубочек в глубь волокна и при помощи особого, пока еще полностью не изученного, механизма вызывает освобождение ионов Са2+ из боковых цистерн. Боковые цистерны расположены таким образом, что освободившиеся ионы Са2+ попадают непосредственно в ту область, где происходит образование актомиозина.
Как отмечалось, начало мышечного сокращения приурочено к первой трети восходящего колена потенциала действия, а именно к моменту, когда внутренний потенциал волокна возрастает с исходных —90 мВ до примерно —50 мВ. Этот потенциал является пороговым для возникновения механического ответа. Предполагают, что именно при достижении указанного уровня деполяризации концентрация свободных ионов Са2+ в миофибрилле достигает критической величины, необходимой для начала взаимодействия актиновых и миозиновых нитей.
Процесс освобождения Са2+ прекращается после окончания пика потенциала действия. Тем не менее сокращение продолжает нарастать до тех пор, пока активация кальциевого насоса ретикулума не вызовет снижения концентрации ионов Са2+ в миоплазме. Тогда сокращение сменяется расслаблением.
Таким образом, последовательность событий, ведущих к сокращению, а затем расслаблению мышечного волокна, представляется в следующем виде: раздражение —>- возникновение потенциала действия -> проведение его вдоль клеточной мембраны и в глубь волокна по трубочкам —> освобождение Са2+ из боковых цистерн саркоплазматического ретикулума и диффузия его к миофибриллам —> взаимодействие («скольжение») актиновых и миозиновых нитей, приводящее к укорочению миофибриллы —> активация кальциевого насоса —> снижение концентрации свободных ионов Ca2+ в саркоплазме —> расслабление миофибрилл.