

Критический уровень деполяризации
Регистрация изменений мембранного потенциала при внутриклеточном раздражении нервного или мышечного волокна показала, что потенциал действия возникает в тот момент, когда деполяризация мембраны достигает критического уровня. Этот критический уровень деполяризации не зависит от характера примененного стимула, расстояния между электродами и т. п., а определяется исключительно свойствами самой мембраны.
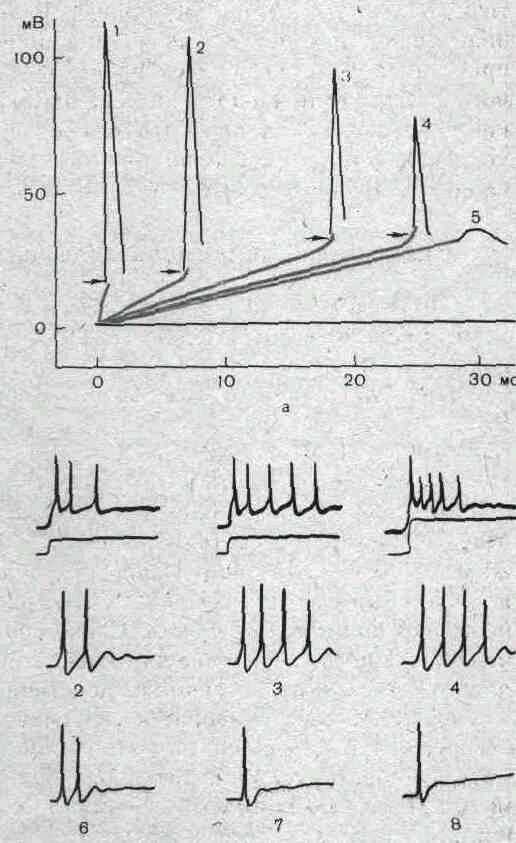
Рис.
14.
Изменение мембранного потенциала
до критического уровня деполяризации
мембраны при действии раздражающего
тока разной силы и длительности.
Критический
уровень показан пунктиром. Внизу
—
раздражающие стимулы, при воздействии
которых получены ответы А, Б и В.
Локальный ответ
В механизме критической деполяризации мембраны наряду с пассивными существенную роль играют активные подпороговые изменения мембранного потенциала, проявляющиеся в форме так называемого локального ответа.

Первые признаки локального ответа появляются при действии стимулов, составляющих 50—75 % от пороговой величины. По мере дальнейшего усиления раздражающего тока локальный ответ увеличивается, и в момент, когда деполяризация мембраны, обусловленная суммой катэлектротонического потенциала и локального ответа, достигает критического уровня, возникает потенциал действия (рис. 15).
Локальный ответ, так же как и потенциал действия, обусловлен повышением натриевой проницаемости мембраны. Однако при подпороговом стимуле это начальное повышение натриевой проницаемости недостаточно велико, чтобы вызвать быструю регенеративную деполяризацию мембраны. Развитие деполяризации тормозится процессами инактивации натриевых и активации калиевых каналов. Поэтому рост локального ответа приостанавливается, а затем происходит реполяризация мембраны. Амплитуда локального ответа увеличивается по мере приближения силы стимула к порогу, и при достижении последнего локальный ответ перерастает в потенциал действия, поскольку скорость увеличения натриевой проницаемости мембраны начинает превышать скорость роста калиевой проницаемости.
Зависимость пороговой силы раздражителя от его длительности
Пороговая сила любого стимула в определенных пределах находится в обратной зависимости от его длительности. Особенно четко эта зависимость проявляется при использовании в качестве раздражителя прямоугольных импульсов постоянного тока.
Представленная на рис. 16 кривая называется кривой силы—длительности, или силы — времени. Она была изучена при исследовании различных нервов и мышц Гоорве-гом (1892), Вейсом (1901) и Лапиком (1909).
По этой кривой прежде всего можно судить о том, что ток ниже некоторой минимальной силы или напряжения не вызывает возбуждения, как бы длительно он ни действовал. Минимальная сила постоянного тока, способная вызвать возбуждение (порог раздражения), названа Лапиком реобазой (ордината ОА). Наименьшее время (отрезок ОС), в течение которого должен действовать раздражающий стимул, величиной в одну реобазу называют полезным временем. Слово «полезное» здесь применено с целью подчеркнуть, что дальнейшее увеличение длительности действия тока не имеет значения (бесполезно) для возникновения потенциала действия.
Усиление тока приводит к укорочению минимального времени раздражения, но не беспредельно. Как видно на рис. 16, при очень коротких стимулах кривая силы — времени становится параллельной оси ординат. Это означает, что при таких кратковременных раздражениях возбуждения не возникает, как бы ни была велика сила раздражителя. Поэтому, кроме полезного времени, в качестве времени константы раздраже-

В настоящее время показано, что величина хронаксии зависит от RC мембраны и временной константы процесса активации натриевых (или кальциевых) каналов. Точное измерение величин реобазы или хронаксии возможно только в опытах на одиночных возбудимых клетках. При раздражении целой мышцы или нерва (особенно через кожу, как это делается при исследованиях возбудимости, проводимых на человеке) ветвление тока и поляризация окружающих тканей вносят очень большие искажения в измеряемые величины. Все же в некоторых случаях использование хро-наксиметрии оказалось полезным в неврологической практике: с ее помощью удается установить наличие органического поражения (перерождения) двигательного нерва. Дело в том, что электрический ток, приложенный к мышце, проходит и через находящиеся в ней нервные волокна и их окончания. Величины реобазы и хронаксии нервных волокон значительно меньше соответствующих величин мышечных волокон, поэтому при пороговых силах тока возбуждение прежде всего возникает в нервных волокнах и от них передается на мышцу. Из этого следует, что при измерении хронаксии мышцы фактически получают значение хронаксии пннервирующих ее нервных волокон. Если нерв поврежден или произошла-гибель соответствующих мотонейронов в спинном мозге (как это, например, имеет место при полиомиелите), то нервные волокна перерождаются, тогда раздражающий стимул выявляет хронаксию собственно мышечных волокон, которая имеет большую продолжительность.