

2.2 Класс Reptilia. Пресмыкающиеся (Рептилии)
Ранние рептилии имели общие черты строения с земноводными и парарептилиями, более позние – с птицами и млекопитающими (появление теплокровности, вторичного неба, четырехкамерного сердца, перьев, шерсти и пр.).
От парарептилий рептилии отличаются, прежде всего, строение черепа: сзади глазниц он не является сплошным, а несет одно или два отверстия – височные окна (ямы).
От земноводных рептилии отличаются следующими основными признаками:
1. Размножением яйцами, имеющими кроме зародышевой оболочки также твердую скорлупу, что в совокупности явилось исключительно важным приспособлением позвоночных к наземной жизни. Из яйца появляется молодое животное, которое отличается от взрослого только размерами.
2. Развитием легочного дыхания, что привело к необходимости защитить голую кожу животного покрытием (роговые чешуи, костные пластинки). Это явилось вторым важным шагом на пути приспособления к сухопутной жизни.
3. Разнообразием условий существования (земля, вода, воздух).
4. Более совершенными и сложными кровообращением и выделительной системой. В процессе дальнейшего развития центральной нервной системы появилась кора мозга в больших полушариях.
5. Значительными изменениями в строении скелета. В височной области появляются височные впадины, служащие вместилищем челюстной мускулатуры (более сильной, чем у амфибий). Позвоночник приобретает четко выраженную дифференцировку: в частности, обособляется шейный отдел (у земноводных отсутствовал). Это дало животному свободу движений. В теле позвонков постепенно исчезает отверстие для хорды. Парные конечности стали разнообразнее и совершеннее: развились суставные поверхности, конечности удлинились. Это позволило поднять тело высоко, не волоча его по земле, что было необходимо для быстрого передвижения.
Вместе с тем у рептилий сохранились и ряд примитивных признаков: плохо развиты органы чувств (кроме зрения), слуховой аппарат (за небольшим исключением) состоит из одной косточки, т.е. как у амфибий, зубы у многих не дифференцированы и т.д. Как и земноводные, большинство пресмыкающихся являются холоднокровными животными с непостоянной температурой тела. И те, и другие живут обычно в теплых широтах.
Рептилии появляются в середине карбона, в мезозое широко распространены, к концу мезозоя большая часть их вымирает. Современные представители класса – змеи, ящерицы, крокодилы, гаттерии и др. В ископаемом состоянии встречаются как полные скелеты рептилий, так и их части. Довольно часты следы лап рептилий. Их можно найти в местах захоронения скелетов животных, или отдельно. Следы очень разнообразны по облику, размерам и сохранности. Обычны копролиты. Наиболее крупные скопления остатков рептилий наблюдаются в области дельт, т.е. там, где были самые благоприятные условия для захоронения: концентрация трупов течением и их быстрое занесение осадком. В озерах же, наоборот, встречаются обычно отдельные скелеты и их части.
С биологической точки зрения значение пресмыкающихся и парарептилий в истории развития органического мира нашей планеты велико: именно они являются непосредственным промежуточным звеном между амфибиями, птицами и млекопитающими. Родственные связи прослеживаются через котилозавров с амфибиями, через архозавров – с птицами, через зверообразных – с млекопитающими.
Геологическое значение рептилий также велико. Сухопутные рептилии (котилозавры, зверообразные, динозавры) широко используются в стратиграфии верхнепалеозойских и мезозойских, в основном континентальных отложений. Остатки морских пресмыкающихся (ихтиозавров, плезиозавров, мозазавров) имеют меньшую ценность из-за их редкости, однако при их обнаружении окаменелости применяют в основном для расчленения верхнеюрских и меловых отложений.
Класс Пресмыкающихся объединяет пять подклассов: Зверообразные –Synapsida (С3-К1), Чешуйчатые – Lepidosauria (Р2-Q), Эвриапсиды – Euryapsida (Т-K), Ихтиозавры или Рыбоящеры – Ichthyopterygia (T-K2), Архозавры – Archosauria (Р2-Q).
Подкласс Зверообразные. Synapsida. Эти рептилии имели синапсидный (одно височное окно) тип черепа. В ходе эволюции приобрели черты млекопитающих: увеличилась зубная кость нижней челюсти, появилась дифференциация зубов (резцы, клыки и коренные зубы), появился шерстистый покров и теплокровность и т.д. Череп зверообразных был высоким и узким. Конечности не отличались по длине или задние были длиннее передних. Многие имели почти прямую постановку конечностей. Большинство синапсид - хищники. Жили со среднего карбона – по среднюю юру.
В конце карбона они возникли от котилозавров, в свою очередь, их представители в триасе дали начало млекопитающим. Примером зверообразных является иностранцевия - Inostrancevia (рис. 30, 31), жившая в перми. Рептилия имела дифференцированные зубы: резцы, заклыковые зубы, мощные клыки, а на пальцах – большие когти. Животное достигало 3 м в длину при длине черепа 50 см.
 Рис.
26. Реконструкция рептилии подкласса
зверообразных – иностранцевии
(Inostrancevia,
P2)
[1]
Рис.
26. Реконструкция рептилии подкласса
зверообразных – иностранцевии
(Inostrancevia,
P2)
[1]
Интересна также двиния (Dvinia) из поздней перми – мелкое всеядное животное с шерстистым покровом, похожее на крысу (рис. 27). Двинии относятся к последнему и самому продвинутому подотряду зверообразных – цинодонтам – прямым предкам млекопитающих. Они уже были полностью покрыты шерстью и мало отличались от первых триасовых млекопитающих. Есть мнение, что современные утконос и ехидна – на самом деле цинодонты, сохранившиеся до наших дней.

Рис. 27. Dvinia – представитель зверообразных –
потомков млекопитающих [45]
Характерные представители зверообразных: иностранцевия – Inostrancevia (P2-3), двиния – Dvinia (P3).
Подкласс Lepidosauria. Чешуйчатые. Это примитивные ящерицеобразные пресмыкающиеся (чешуйчатые ящеры), известные с перми. В палеозое они имели диапсидный тип черепа (две височные дуги и два окна). В течение мезозоя и кайнозоя дуги и окна частично или полностью исчезли.
Чашуйчатые имели парные конечности в основном умеренной длины, но у змей и некоторых ящериц они исчезают, хотя рудименты поясов конечностей могут сохраняться. У некоторых водных форм конечности становятся ластовидными. У лепидозавров сохраняется обычный для ранних четвероногих примитивный тип передвижения с волнообразным изгибанием туловища.
Современные представители подкласса – змеи, ящерицы, хамелеоны, гаттерии. Змеи обособились от ящериц в конце мела.
Лепидозавры жили в разных услових (морях, болотах, озерах, на суше и т.д.), поэтому их остатки встречают в разных породах. Стратиграфическое значение лепидозавров невелико. Но те из них, которые имеют узкий диапазон распространения, например, мозозавры, с успехом используются для расчленения мезозойских отложения.
Вымершие чешуйчатые представлены наземными ящерицеобразными эозухиями и хищными морскими ящерицами – мозозаврами. Они имеют диапсидный тип черепа и являются переходной группой между парарептилиями – котилозаврами и прогрессивными рептилиями – архозаврами.
Эозухии (рис. 28) внешне были похожи на ящериц или крокодилов. Первые имели небольшие размеры – до 50 см, крокодилообразные могли достигать 7,5 м. От настоящих ящериц и крокодилов отличались строением черепа. Они появились в перми, в триасе дали начало другим чешуйчатым: клювоголовым и ящерицам. Поздний карбон – ранний триас.


Рис. 28. Эозухии: а – скелет [6], б – реконструкция [39]
Мозозавры (рис. 29) – морские ящерицы, жившие в позднем мелу, имели удлиненное тело до 15 м в длину, крупную голову и две пары пятипалых конечностей – ласт, хвост превращен в плавник, который являлся главным органом движения.

-
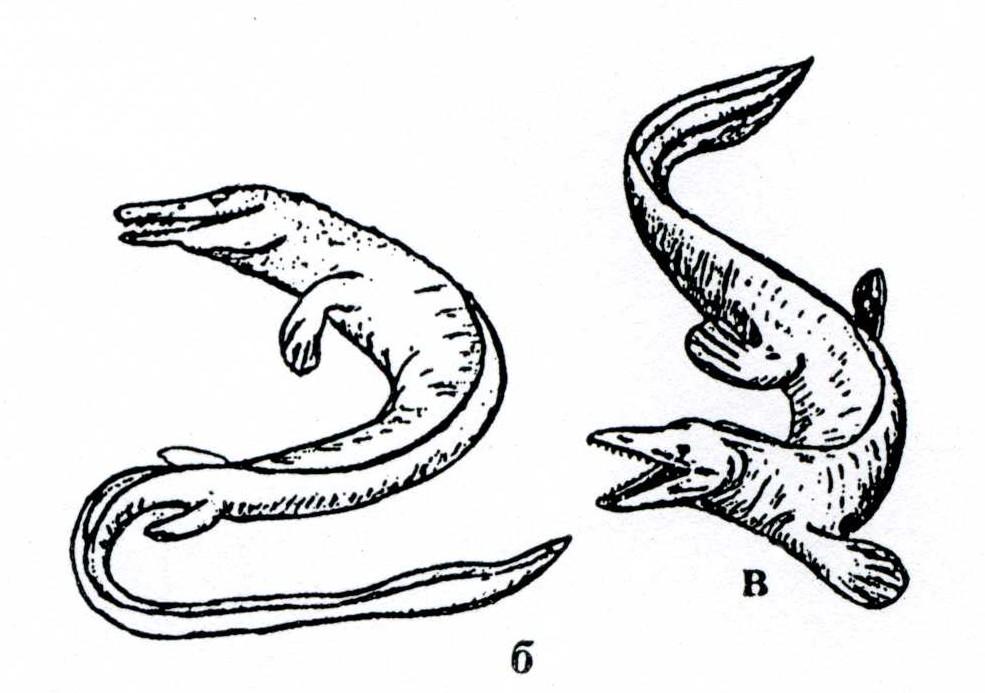
Рис. 29. Мозозавры – морские ящерицы: а – скелет [6], б-в – реконструкция животных
Подкласс Эвриапсиды (Euryapsida) или Synaptosauria. Синаптозавры.. Эти пресмыкающиеся вели водный (морской) образ жизни и все являются плотоядними. Они имели обычно крупное, широкое (боченковидное) туловище, короткий хвост и часто длинную шею. Длина тела достигала 16 м. Тип черепа эвриапсидный, характеризующийся одной верхней височной ямой. Конечности – ластовидные.
Синаптозавры представлены двумя отрядами. Представители одного из них (плезиозавры, плиозавры и др.) имели конические острые зубы (T-K), другого – уплощенные зубы (плакодонты) (T2-J1).
Плезиозавры (рис. 30) – животные с маленькой головой, длинной, гибкой шеей и коротким хвостом. Длина до 16 м. Это активные хищники. Обитали вблизи берега, питались рыбой. Классические местонахождения плезиозавров находятся в Англии. Триас-мел.

Рис. 30. Плезиозавр – муренозавр (Muraenosaurus, J3) [6]

Рис. 31. Плезиозавры (реконструкция) [14]
Плиозавры (рис. 32) – хищники огромных размеров с короткой шеей и крупной головой (череп до 4 м в длину), с сильными коническими зубами и хвостом разной длины. Длина тела до 15 м. Обитатели открытых морей в триасе и мелу. Принадлежали к числу опаснейших морских хищников своего времени.


Рис. 32. Плиозавры: а – череп плиозавра сбоку [6], б – реконструкция [40].
Плакодонты (рис. 33) – морские пресмыкающиеся, часть которых похожа на черепах. Они имели уплощенное (иногда удлиненное) тело, защищенное костными щитками, иногда образующими полный панцирь. Шея короткая и толстая. Конечности ластовидные. Те плакодонты, которые имели острые зубы, являлись малоподвижными хищниками морского мелководья, они питались моллюсками и брахиоподами. Те, у которых вместо зубов имеется клюв, питались планктоном и водорослями. Длина плакодонтов достигала 2,5 м. Могли некоторое время проводить на суше. Средний триас – ранняя юра.
Есть мнение, что плезиозавры, плиозавры и плакодонты возникли от котилозавров. Согласно другой точке зрения, синаптозавры берут свое начало от примитивных диапсид, вероятно, близким эозухиям (см. ниже).


Рис. 33. Плакодонты: а – скелет (Placochelys, T3) [6], б – реконструкция [20]
Остатки синаптозавров встречаются редко, поэтому стратиграфическое значение их невелико. Тем не менее, плезиозавры и плиозавры входят в состав типовых комплексов позвоночных верхнего триаса-нижнего мела и используются для расчленения и определения возраста отложений этого уровня. Плакодонты, имеющие узкое распространение, используются как руководящие формы для триаса.
Подкласс Ихтиозавры, или рыбоящеры. Ichthyopterygia. Подкласс объединил морских рептилий, утративших связь с сушей. Эти животные имели рыбообразное тело, мягкий спинной плавник и жесткие хвостовые плавники, парные конечности - ласты (рис. 34). Ласты – это видоизмененные конечности первых четвероногих. Длина тела некоторых форм достигала 23 м. Череп с длинной мордой и одной небольшой височной ямой (эвриапсидный тип). Зубы хорошо развиты, они конические, однорядные, лабиринтовые. Посредине хвост в той или иной степени изогнут вниз: у триасовых форм – незначительно, у юрских и меловых - очень резко. Чешуйчатый покров не установлен, но в коже была развита система роговых пластин. Кожа для лучшего скольжения в воде была, возможно, покрыта слизью.

Рис. 34. Ихтиозавр (реконструкция) [19].
У типичного ихтиозавра были очень большие глаза (до 20 см в диаметре), защищённые костяным кольцом, говорящим о том, что охотились они ночью. Ихтиозавры не имели жабр и дышали воздухом. Они не могли выходить на землю, поэтому перешли к живорождению, о чем свидетельствуют находки скелетов молодых особей внутри скелетов взрослых.
Ихтиозавры внешне похожи на рыб и дельфинов, к рыбам их первоначально и относили. Многие из ихтиозавров были хищниками, они питались моллюсками, в том числе белемнитами, рыбой, мелкими рептилиями, а также своими же сородичами.
Ихтиозавры известны с верхней части нижнего триаса, их расцвет приходится на раннюю юру, в позднем мелу они вымирают. Ихтиозавры, как и другие водные рептилии, имеют меньшую стратиграфическую ценность по сравнению с сухопутными рептилиями из-за их относительной малочисленности, тем не менее, они используются для расчленения юрских и меловых отложений.
Ихтиозавры захоронены в сланцах, мергелях, глинах, доломитах совместно с белемнитами, иглокожими, рыбами и др. Многочисленны как находки отдельных костей этих животных, так и целых скелетов, иногда с отпечатками мягких тканей. Скелеты могут быть заключены в известковые конкреции. Основные местонахождения остатков образовались на месте мелководных бухт.
Характерный представитель: ихтиозавр - Ichthyosaurus(MZ).
Подкласс Архозавры, или древние ящерицы. Archosauria. Самая многочисленная группа рептилий с диапсидным типом (две височные впадины и часто предглазничное отверстие) черепа. По способу размножения архозавры были яйцекладущими животными. Архозавры произошли от чешуйчатых – эозухий в перми. В настоящее время они представлены крокодилами.
В составе подкласса выделяются надотряды и отряды, в том числе вымершие надотряды текодонтов, динозавров и птерозавров.
Надотряд Текодонты. Thecodontia – сборная группа, включающая первых архозавров, живших в перми и триасе. В последнее время ее представителей включают в группу круротарзов.
Внешне текодонты напоминали ящериц, динозавров и крокодилов. Передвигались на четырех или двух ногах. Они имели длинный узкий череп и текодонтные зубы. Это зубы, которые расположены в особых ячейках по краям челюстей - теках, которые снижали риск потерять их в процессе питания. Передние конечности у текодонтов были короче, чем задние.
Текодонты разделяются на два отряда: фитозавров (рис. 35) и псевдозухий (рис. 36). Фитозавры – крупные (до 5-6 м) крокодилообразные текодонты с удлиненным рылом и кожным панцирем, жившие в пресноводных бассейнах триаса и питавшиеся рыбой. Псевдозухии (рис. 36) - некрупные подвижные наземные животные ящерицеобразной и крокодилообразной формы с панцирем или без него. Среди последних присутствовали хищные, плотоядные и насекомоядные рептилии. Жили с перми по триас.


Рис. 35. Фитозавры: а - скелет [6], б – реконструкция [35]
Текодонты дали начало всем остальным группам архозавров: разнообразным динозаврам, летающим ящерам и крокодилам.
Представители надотряда используются в стратиграфии в основном триасовых отложений.

Рис. 36. Псевдозухия (реконструкция) [15]
Надотряд динозавров. Dinosauria (страшная ящерица). Это – самая многочисленная, господствующая в мезозое, группа рептилий. Жили с позднего триаса по поздний мел. Объединяет разнообразных по морфологии, размерам и образу жизни животных. Здесь есть небольшие рептилии величиной с кошку, рептилии средних размеров (большинство) и крупные - до 20-35 м (редко) в длину, двуногие и четвероногие, хищники и питавшиеся растениями, несшие костные панцири, «голые» и оперенные.
Выделяются два отряда динозавров: ящеротазовые и птицетазовые.
Отряд Ящеротазовые динозавры. Делятся на две группы: звероногие (тероподы) и ящероногие (завроподы). Появились в триасе, расцвет группы приходится на юру и ранний мел, в конце мела вымирают.
Т е р о п о д ы объединили двуногих животных с укороченными передними конечностями, которые передвигались на задних лапах, опираясь на сильный хвост. Среди них были хищники и растительноядные. Размеры достигали 14 м в длину. Некоторые тероподы имели перьевой покров.
Характерный представитель группы – тираннозавр (рис. 37). Известные в настоящее время его мелкие предки (например, дилонг) были оперены тонкими волосовидными перьями. У самого тираннозавра перьев могло и не быть (известные отпечатки кожи бедра тираннозавра несут типичный для динозавров рисунок из полигональных чешуй).
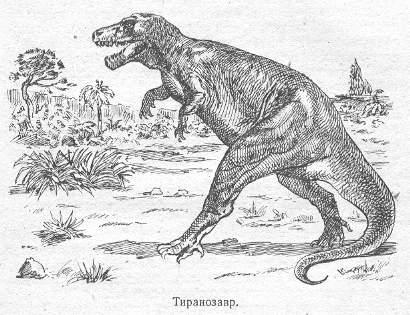
Рис. 37. Звероногий динозавр (теропод) – тираннозавр (Tirannosaurus, K2), реконструкция внешнего вида [24]
Большинство ученых с недавних пор стали относить к тероподам современных птиц (в качестве одного из подотрядов). Среди терапод можно найти формы, покрытые многочисленными пупырышками, волокнистым пухом или обладающих настоящими перьями.
Первым динозавром, у которого была установлена прижизненная окраска перьев, является синозавроптерикс (Sinosauropteryx), живший в раннем мелу (рис. 38). Ученым удалось выявить в останках его перьевого покрова окаменелые меланосомы — клеточные органеллы, в которых находятся молекулы меланина. На основе полученных исследований был сделан вывод, что синозавроптерикс имел яркое оперение: хвост был окрашен оранжево-белыми полосами, верхняя часть туловища имела рыжий цвет, живот был светлым, а ноги темными. Длина динозавра около 1 м.

Рис. 38. Теропод синозавроптерикс (Sinosauropteryx, К1) [30]
З а у р о п о д ы (рис. 39, 40, 41, 42) относились к растительноядным организмам. Они передвигались на четырех лапах. Размеры их головного мозга были очень маленькими.

Рис. 39. Ящероногий динозавр – диплодок (Diplodocus, J3).

Рис. 40. Кладка яиц крупного растительноядного динозавра-зауропода мелового периода (фото Т.Ю.Дрозд).
Зауроподы были самыми крупными животными, жившие когда-либо на нашей планете. Так, диплодоки и им подобные динозавры имели длину тела до 36 м и вес до 80 тонн. Считается, что они вели наземный, сухопутный образ жизни (как слоны) и перемещались по зарослям и степям от водоема к водоему.
Характерные представители: диплодоки, брахиозавры. Они, при общем внешнем сходстве, отличаются многими признаками. В частности, у диплодоков задние конечности длиннее передних, а у брахиозавров, наоборот, - передние существенно длиннее задних. Кроме того, у брахиозавров гребень на голове.

Рис. 41. На переднем плане – группа диплодоков [17].

Рис. 42. Брахиозавр (Brachiosaurus) [37]
Отряд Птицетазовые динозавры. Все рептилии этого отряда являются растительноядными. Они имели разные размеры и морфологию, многие имели защитные приспособления: панцирь, рога, щиты и др. Появляются в начале мезозоя, а к концу мела вымирают. Выделяются четыре группы этих динозавров: птиценогие динозавры, стегозавры, панцирные и рогатые динозавры.
Птиценогие динозавры (игуанодонтиды) – высокие динозавры, ходившие на двух задних конечностях. Они жили на суше и в неглубоких водоемах. Один из их представителей, игуанодон (рис. 43), представлял собой крупное животное с длиной тела до 5-10 м и черепом похожим на лошадиный. Зубы напоминали ряд зубов млекопитающих. Передние конечности игуанодонов были короткими, пятипалыми, задние конечности были трехпалые с копытами. Обитал стадами вблизи водоемов. Ранний мел.
|
|
|


Рис. 43. Птиценогий динозавр игуанодон (Iguanodon, K1): а – скелет [6] [основы], б – реконструкция животного [31]
Стегозавры (рис. 44) первоначально передвигались на задних конечностях, а позже перешли к четвероногому хождению. Они имели большое туловище, маленькую голову и крупный хвост. Длина тела до 6 м.
|
|
|


Рис. 44. Птицетазовый динозавр - стегозавр (Stegosaurus, J3):
а- скелет [6], б – реконструкция [42]
В качестве защитного приспособления на их спинах имелись костные пластинки размером до 1 м, а на конце хвоста – острые шипы. Распространение стегозавров – юра-ранний мел.
Панцирные динозавры (анкилозавры) (рис. 45) были крупными животными с широким уплощенным телом (длина до 5 м), покрытым массивным панцирем из костных пластин. Стратиграфическое распространение их узкое – мел.

Рис. 45. Панцирный динозавр – анкилозавр (Ankylosaurus, K2) [44].
В позднем мелу обитали рогатые динозавры(цератопсы) – крупные (до 7 м в длину) грузные рептилии, ходившие на четырех ногах. Характерная их особенность – наличие на черепе костного «воротника», который защищал область шеи. На конце морды и над глазами располагались рога. Типичный представитель группы –трицератопс(рис. 46).
|
|
|


Рис. 46. Рогатый динозавр трицератопс (Triceratops, K2): а – скелет [6], б – реконструкция [41]
Надотряд Pterosauria. Птерозавры (крылатые ящеры) – летающие животные, внешне напоминающие птиц (рис. 47). Четырехпалые с сильно удлиненным четвертым пальцем передние конечности птерозавров были преобразованы в крылья. Между конечностями и боками тела была натянута летательная перепонка. Она представляла собой вытянутую складку кожи, которая крепилась на очень длинном четвертом пальце. Остальные три пальца короткие и оканчивались когтями. Птерозавры имели облегченные кости с воздушными полостями и хорошо развитую грудину с мощной мускулатурой. Челюсти – вытянутые, снабженные нередко зубами. Хвосты - разной длины.
Птерозавры являлись теплокровными рептилиями. У молодых особей имелся плотный «шерстистый» (волосовидный) покров.
По типу питания птерозавры насекомоядные, рыбоядные, некоторые питались трупами или планктоном, а также беспозвоночными.
Птерозавры произошли от текодонтов. Жили с позднего триаса до конца мела. Стратиграфическое значение их невелико.
Характерными представителями надотряда являются рамфоринхусы, птеродактили и птеранодоны. Первые (рис. 47а, 48) были размером с небольшую птицу, имели зубы и длинный хвост, заканчивающийся кисточкой. Летательные пальцы не складывались.

Рис. 47. Птерозавры, или крылатые ящеры:
а – длиннохвостый рамфоринхус (Rhamphorhinchus, J),
б – гигантский короткохвостый птеранодон (Pteranodon, K) [1]
Размеры птеродактилей - также небольшие, их клюв был снабжен зубами, но хвост был коротким или отсутствовал (рис. 50). Птеронодоны являлись беззубыми ящерами с коротким хвостом и высоким гребнем в верхней части черепа, имели большие размеры (размах крыльев достигал 16 м) (рис.47б, 49). У птеродактилей и птеранодонов летательные пальцы складывались.

Рис. 48. Рамфоринхус (Rhamphorhinchus, J) – реконструкция [16].

Рис.49. Птеранодон (Pteranodon, K) [28]
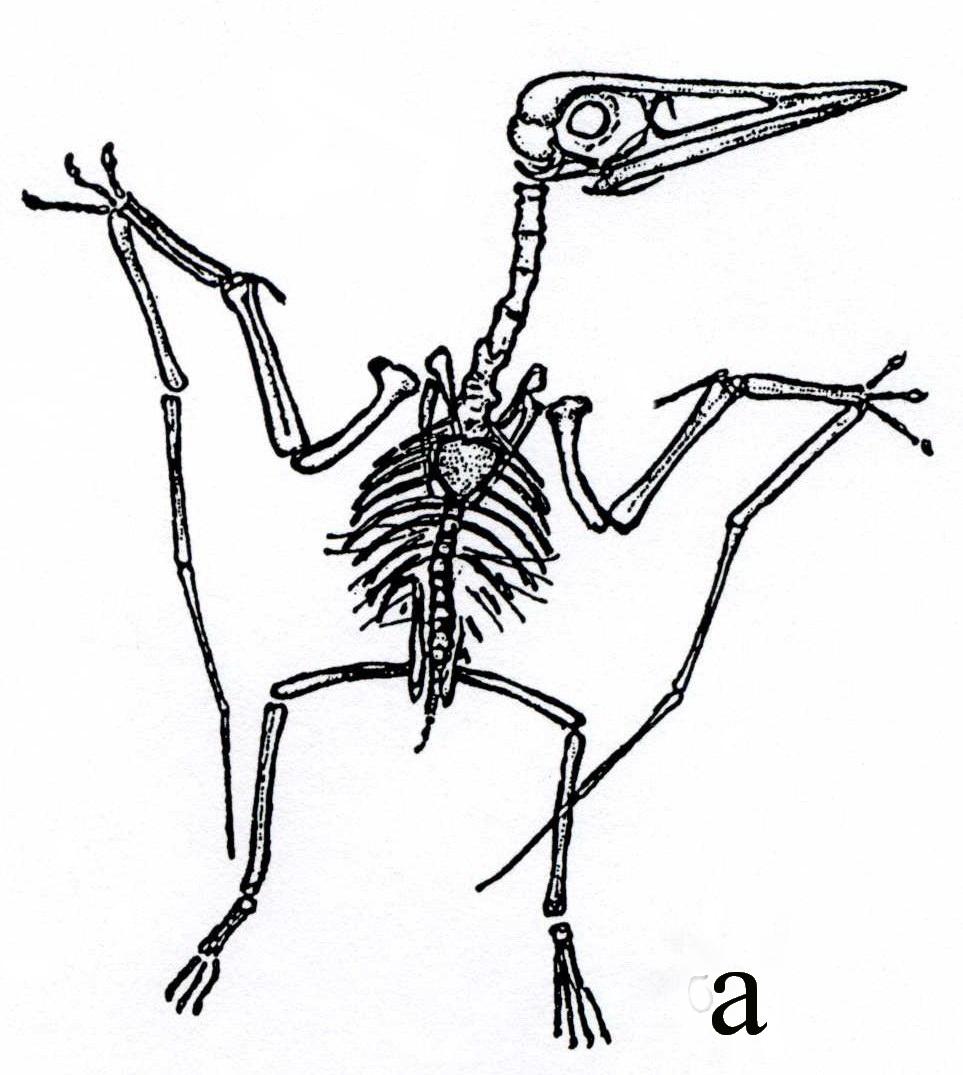

Рис. 50. Птеродактиль (J): а - скелет, длина – 12 см [6], б – реконструкция внешнего вида [18]
Подкласс Crocodilia. Крокодилы. Т2-ныне. (рис. 51). Тело этих рептилий покрыто роговыми щитками, под которыми расположены костные пластинки. Туловище приплюснуто, хвост – длинный, задние конечности длиннее передних.
Крокодилы произошли от текодонтов в триасе, являются самыми организованными из современных пресмыкающихся. Сейчас населяют пресные и опресненные водоемы тропиков. Все крокодилы – хищники. Морские крокодилы используются в стратиграфии юры, пресноводные – верхнего мела.

Рис. 51. Телеозавры - ископаемые морские крокодилы юрского периода [33]