

4. Хлоропласти. Функції та хімічний склад.
Розмір хлоропластів коливається від 4 до 10 мкм. Число хлоропластів зазвичай становить від 20 до 100 на клітину. Хімічний склад хлоропластів досить складний і може бути охарактеризований такими середніми даними (% на суху масу): білок - 35-55; ліпіди - 20-30; вуглеводи - 10; РНК - 2-3; ДНК - до 0,5; хлорофіл - 9; каротиноїди - 4,5. Важливо відзначити, що багато білки хлоропластів володіють ферментативної активністю. Дійсно, в хлоропластах зосереджені всі ферменти, що беруть участь в процесі фотосинтезу (окисно-відновні, синтетази гідролази). В даний час доведено, що в хлоропластах, так само як і в мітохондріях, мається своя білоксинтезуюча система. Багато з ферментів, локалізованих в хлоропластах, є двокомпонентними. У багатьох випадках простетическая група ферментів - це різні вітаміни. В хлоропластах зосереджено багато вітаміни та їх похідні (вітаміни групи В, К, Е, D). В хлоропластах знаходиться 80% Fe, 70% Zn, близько 50% Сі від усієї кількості цих елементів в листі. Внутрішня будова хлоропластів, їх ультраструктура були розкриті з використанням електронного мікроскопа. Виявилося, що хлоропласти оточені подвійною мембраною. Товщина кожної мембрани 7,5-10 нм, відстань між ними 10-30 нм. Внутрішній простір хлоропластів заповнене безбарвним вмістом - стромой і пронизане мембранами (ламелами). Ламелли, з'єднані один з одним, утворюють хіба бульбашки-тилакоиди (грец. «Тілакоідес» - мешковидний). В хлоропластах містяться тилакоиди двох типів. Короткі тилакоиди зібрані в пачки і розташовані один над одним, нагадуючи стопку монет. Ці стопки називаються гранами, а складові їх тилакоиди - тілакоїдамі гран. Між гранами паралельно один одному розташовуються довгі тилакоиди. Складові їх ламелли отримали назву - тилакоиди строми. Між окремими тілакоїдамі в стопках гран є вузькі щілини. Тілакоідние мембрани містять велику кількість білків, що у фотосинтезі. У складі інтегральних мембранних білків є багато гідрофобних амінокислот. Це створює безводну середовище і робить мембрани стабільніше. Багато білки тілакоідних мембран побудовані у вигляді векторів і межують з одного боку з стромою, а з іншого контактують з внутрішнім простором тилакоида.
Щодо
зв'язку між ламелами гран і ламелами
строми є різні точки зору. Т. Вейер
запропонував гранулярності-ґратчасту
модель, згідно з якою внутрішні простори
всіх тилакоидов з'єднані між собою.
Таким чином, в хлоропластах мається як
би два роздільних простору - внутрішнє
(всередині тилакоидов) і зовнішнє (поза
тилакоидов). У більшості водоростей
гран немає, а ламелли зібрані в групи
(пачки) по 2-8 штук. Не у всіх випадках і
у вищих рослин хлоропласти мають
гранальную структуру. Так, в листі
кукурудзи є виду хлоропластів. В клітинах
мезофилла містяться дрібні хлоропласти
гранального будови. В клітинах обкладки,
оточуючих листові судинні пучки,
хлоропласти крупні і гран не містять.
В стромі хлоропластів знаходяться нитки
ДНК, рибосоми, крохмальні
зерна. Основний
фермент, що забезпечує засвоєння
вуглекислого газу, -
рибузолобифосфаткарбоксилазаоксигеназа
(скорочено РБФ - карбоксілазаоксігеназа)
також розташований в стромі. Зелений
пігмент хлорофіл у вигляді комплексу
з білками (пігмент-білкові комплекси)
зосереджений головним чином в Тилакоїди
гран і частково в Тилакоїди строми. В
мембранах тилакоїдів гран локалізована
фермент, що каталізує синтез АТФ
(АТФ-синтаза). Цей фермент пов'язаний з
білком, розташованим в самій мембрані.5.
Мітохондрії. Хімічний склад.Мітохондрії
- «силові» станції клітини, в них
локалізована велика частина реакцій
дихання (аеробне фаза). В мітохондріях
відбувається акумуляція енергії дихання
в аденозинтрифосфату (АТФ). Енергія,
запасна в АТФ, служить основним джерелом
для фізіологічної діяльності клітини.
Мітохондрії зазвичай мають видовжену
палочковидную форму довжиною 4-7 мкм і
діаметром 0,5-2 мкм. Число мітохондрій в
клітині може бути різним від 500 до 1000 і
залежить від ролі даного органу в
процесах енергетичного обміну. Проте
в деяких організмах (дріжджах) мається
лише одна гігантська мітохондрія.
Основний
фермент, що забезпечує засвоєння
вуглекислого газу, -
рибузолобифосфаткарбоксилазаоксигеназа
(скорочено РБФ - карбоксілазаоксігеназа)
також розташований в стромі. Зелений
пігмент хлорофіл у вигляді комплексу
з білками (пігмент-білкові комплекси)
зосереджений головним чином в Тилакоїди
гран і частково в Тилакоїди строми. В
мембранах тилакоїдів гран локалізована
фермент, що каталізує синтез АТФ
(АТФ-синтаза). Цей фермент пов'язаний з
білком, розташованим в самій мембрані.5.
Мітохондрії. Хімічний склад.Мітохондрії
- «силові» станції клітини, в них
локалізована велика частина реакцій
дихання (аеробне фаза). В мітохондріях
відбувається акумуляція енергії дихання
в аденозинтрифосфату (АТФ). Енергія,
запасна в АТФ, служить основним джерелом
для фізіологічної діяльності клітини.
Мітохондрії зазвичай мають видовжену
палочковидную форму довжиною 4-7 мкм і
діаметром 0,5-2 мкм. Число мітохондрій в
клітині може бути різним від 500 до 1000 і
залежить від ролі даного органу в
процесах енергетичного обміну. Проте
в деяких організмах (дріжджах) мається
лише одна гігантська мітохондрія. Хімічний
склад мітохондрій дещо коливається. В
основному це білково-ліпідні органели.
Вміст білка в них становить 60-65%, причому
структурні та ферментативні білки
містяться приблизно в рівній пропорції,
а також близько 30% ліпідів. Дуже важливо,
що мітохондрії містять нуклеїнові
кислоти: РНК - 1% і ДНК -0,5%. В мітохондріях
мається не тільки ДНК, але і вся система
синтезу білка, в тому числі рибосоми.
Мітохондрії оточені подвійною мембраною.
Товщина мембран становить 6-10 нм. Мембрани
мітохондрій на 70% складаються з білка.
Фосфоліпіди мембран представлені
фосфатідтілхоліном, фосфатидилетаноламін,
а також специфічними фосфолипидами,
наприклад, кардіоліпіну. Мембрани
мітохондрій не пропускають Н + і служать
бар'єром для їх транспорту. Це найважливіша
властивість лежить в основі утворення
електрохімічного градієнта. Між
мембранами знаходиться заповнену
рідиною перімітохондріальное простір,
рівний 10 нм. Внутрішній простір мітохондрій
заповнює матрикс у вигляді студнеобразной
полужидкой маси. В матриксі зосереджені
ферменти циклу Кребса. Внутрішня мембрана
дає вирости - кристи, розташовані
перпендикулярно поздовжньої осі органели
і перегораживающие весь внутрішній
простір мітохондрій на окремі відсіки.
Однак, оскільки вирости-перегородки
неповні, між цими відсіками зберігається
зв'язок. Мембрани мітохондрій мають
велику міцність і гнучкістю. У внутрішній
мембрані локалізована дихальна ланцюг
(ланцюг переносу електронів). На внутрішній
поверхні внутрішньої мембрани мітохондрій
рівномірно розташовані грибоподібні
частинки. Кожна мітохондрія містить
104-105 таких грибоподібних частинок, які
представляють собою фермент АТФ-синтазу,
каталізують освіту АТФ.Мітохондріі
здатні до руху. Це має велике значення
в житті клітини, так як мітохондрії
пересуваються до тих місць, де йде
посилене споживання енергії. Вони можуть
асоціювати один з одним, як шляхом
тісного зближення, так і за допомогою
сполучних тяжів. Спостерігаються також
контакти мітохондрій з ендоплазматичної
мережею, ядром, хлоропластами. Відомо,
що мітохондрії здатні до набухання, а
при втраті води - до зменшення обсягу.
У зростаючих клітинах в мітохондріальному
матриксе кількість крист зростає - це
корелює із збільшенням інтенсивності
дихання. В процесі дихання ультраструктура
мітохондрій змінюється. У тому випадку,
якщо в мітохондріях протікає активний
процес перетворення енергії окислення
в енергію АТФ, внутрішня частина
мітохондрій стає більш компактною.
Мітохондрії мають свій онтогенез. В
меристематических клітинах можна
спостерігати ініціальні частинки, які
представляють собою округлі освіти,
оточені подвійною мембраною. Діаметр
таких ініціальних частинок становить
50 нм. У міру зростання клітини ініціальні
частки збільшуються в розмірі, подовжуються
і їх внутрішня мембрана утворює вирости,
перпендикулярні осі мітохондрій.
Спочатку утворюються промітохондріі.
Вони ще не досягають остаточного розміру
і мають мало крист. З промітохондріі
утворюються мітохондрії. Сформовані
мітохондрії діляться шляхом перетяжки
або брунькуванням.
Хімічний
склад мітохондрій дещо коливається. В
основному це білково-ліпідні органели.
Вміст білка в них становить 60-65%, причому
структурні та ферментативні білки
містяться приблизно в рівній пропорції,
а також близько 30% ліпідів. Дуже важливо,
що мітохондрії містять нуклеїнові
кислоти: РНК - 1% і ДНК -0,5%. В мітохондріях
мається не тільки ДНК, але і вся система
синтезу білка, в тому числі рибосоми.
Мітохондрії оточені подвійною мембраною.
Товщина мембран становить 6-10 нм. Мембрани
мітохондрій на 70% складаються з білка.
Фосфоліпіди мембран представлені
фосфатідтілхоліном, фосфатидилетаноламін,
а також специфічними фосфолипидами,
наприклад, кардіоліпіну. Мембрани
мітохондрій не пропускають Н + і служать
бар'єром для їх транспорту. Це найважливіша
властивість лежить в основі утворення
електрохімічного градієнта. Між
мембранами знаходиться заповнену
рідиною перімітохондріальное простір,
рівний 10 нм. Внутрішній простір мітохондрій
заповнює матрикс у вигляді студнеобразной
полужидкой маси. В матриксі зосереджені
ферменти циклу Кребса. Внутрішня мембрана
дає вирости - кристи, розташовані
перпендикулярно поздовжньої осі органели
і перегораживающие весь внутрішній
простір мітохондрій на окремі відсіки.
Однак, оскільки вирости-перегородки
неповні, між цими відсіками зберігається
зв'язок. Мембрани мітохондрій мають
велику міцність і гнучкістю. У внутрішній
мембрані локалізована дихальна ланцюг
(ланцюг переносу електронів). На внутрішній
поверхні внутрішньої мембрани мітохондрій
рівномірно розташовані грибоподібні
частинки. Кожна мітохондрія містить
104-105 таких грибоподібних частинок, які
представляють собою фермент АТФ-синтазу,
каталізують освіту АТФ.Мітохондріі
здатні до руху. Це має велике значення
в житті клітини, так як мітохондрії
пересуваються до тих місць, де йде
посилене споживання енергії. Вони можуть
асоціювати один з одним, як шляхом
тісного зближення, так і за допомогою
сполучних тяжів. Спостерігаються також
контакти мітохондрій з ендоплазматичної
мережею, ядром, хлоропластами. Відомо,
що мітохондрії здатні до набухання, а
при втраті води - до зменшення обсягу.
У зростаючих клітинах в мітохондріальному
матриксе кількість крист зростає - це
корелює із збільшенням інтенсивності
дихання. В процесі дихання ультраструктура
мітохондрій змінюється. У тому випадку,
якщо в мітохондріях протікає активний
процес перетворення енергії окислення
в енергію АТФ, внутрішня частина
мітохондрій стає більш компактною.
Мітохондрії мають свій онтогенез. В
меристематических клітинах можна
спостерігати ініціальні частинки, які
представляють собою округлі освіти,
оточені подвійною мембраною. Діаметр
таких ініціальних частинок становить
50 нм. У міру зростання клітини ініціальні
частки збільшуються в розмірі, подовжуються
і їх внутрішня мембрана утворює вирости,
перпендикулярні осі мітохондрій.
Спочатку утворюються промітохондріі.
Вони ще не досягають остаточного розміру
і мають мало крист. З промітохондріі
утворюються мітохондрії. Сформовані
мітохондрії діляться шляхом перетяжки
або брунькуванням.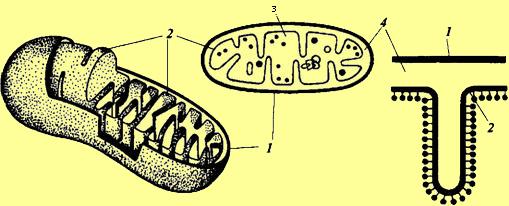 Схема
будови мітохондрії:
Схема
будови мітохондрії:
1 - зовнішня мембрана;
3 - матрикс;
2 - внутрішня мембрана;
4 - перімітохондріальное простір.
Властивості мітохондрій (білки, структура) закодовані частково в ДНК мітохондрій, а частково в ядрі. Так, мітохондріальний геном кодує білки рибосом і частково систему переносників електронотранспортной ланцюга, а в геномі ядра кодована інформація про білках-ферментах циклу Кребса. Зіставлення розмірів мітохондріальної ДНК з числом і розміром мито-хондріальних білків показує, що в ній закладено інформації майже для половини білків. Це і дозволяє вважати мітохондрії, як і хлоропласти, напівавтономними, т. Е. Неповністю залежними від ядра. Вони мають власну ДНК і власну белоксінтезірующую систему, і саме з ними і з пластидами пов'язана так звана цитоплазматическая спадковість. У більшості випадків це спадкування по материнській лінії, так як ініціальні частки мітохондрій локалізовані в яйцеклітині. Таким чином, мітохондрії завжди утворюються від мітохондрій. Широко обговорюється питання, як розглядати мітохондрії і хлоропласти з еволюційної точки зору. Ще в 1921 р російський ботанік Б.М. Козо-Полянський висловив думку, що клітина - це сімбіотрофная система, в якій живе разом кілька організмів. В даний час ендосімбіотіческая теорія походження мітохондрій і хлоропластів є загальноприйнятою. Відповідно до цієї теорії, мітохондрії - це в минулому самостійні організми. На думку Л. Маргеліс (1983), це могли бути еубактеріі, містять ряд дихальних ферментів. На певному етапі еволюції вони проникали в примітивну, що містить ядро, клітку. Виявилося, що ДНК мітохондрій і хлоропластів за своєю структурою різко відрізняється від ядерної ДНК вищих рослин і подібна з бактеріальної ДНК (кільцеве будова, нуклеотидная послідовність). Схожість виявляється і по величині рибосом. Вони дрібніші цитоплазматических рибосом. Синтез білка в мітохондріях, подібно бактериальному, пригнічується антибіотиком хлорамфениколом, який не впливає на синтез білка на рибосомах еукаріот. Крім того, система переносу електронів у бактерій розташована в плазматичної мембрані, що нагадує організацію електронтранспортной ланцюга у внутрішній мітохондріальної мембрані.6. Ядро. Функції та хімічний склад.Сам термін "ядро" вперше був застосований Броуном в 1833 г. Для позначення кулястих постійних структур в клітинах рослин. Пізніше таку ж структуру описали у всіх клітинах вищих організмів. Клітинне ядро звичайно одне на клітину (є приклади багатоядерних клітин), складається з ядерної оболонки, яка відділяє його від цитоплазми, хроматину, ядерця, каріоплазми (або ядерного соку). Ці чотири основні компоненти зустрічаються практично у всіх неделящихся клітинах еукаріотичних одно-і багатоклітинних організмів. Ядра мають звичайно кулясту або яйцеподібну форму; діаметр перших дорівнює приблизно 10 мкм, а довжина друге - 20 мкм. Ядро необхідно для життя клітини, оскільки саме воно регулює всю її активність. Пов'язано це з тим, що ядро несе в собі генетичну (спадкову) інформацію, укладену в ДНК. Ядро мається на клітинах всіх еукаріот за винятком еритроцитів ссавців. У деяких найпростіших є два ядра, але як правило, клітина містить тільки одне ядро. Ядро зазвичай приймає форму кулі або яйця; за розмірами (10-20 мкм) воно є найбільшою з органел. Ядро відмежоване від цитоплазми ядерною оболонкою, яка складається з двох мембран: зовнішньої і внутрішньої, що мають таку ж будову, як і плазматична мембрана. Між ними знаходиться вузький простір, заповнене полужидким речовиною. Через безліч пір в ядерній оболонці здійснюється обмін речовин між ядром і цитоплазмою (зокрема, вихід і-РНК в цитоплазму). Зовнішня мембрана часто буває всіяна рибосомами, синтезуючими білок. Під ядерної оболонкою знаходиться кариоплазма (ядерний сік), до якої надходять речовини з цитоплазми. Каріоплазма містить хроматин - речовина, що несе ДНК, і ядерця. Ядерце - це округла структура всередині ядра, в якій відбувається формування рибосом. Сукупність хромосом, що містяться в хроматині, називають хромосомним набором. Число хромосом в соматичних клітинах диплоидное (2n), на відміну від статевих клітин, що мають гаплоїдний набір хромосом (n). Найважливішою функцією ядра є збереження генетичної інформації. При розподілі клітини ядро також ділиться надвоє, а що знаходиться в ньому ДНК копіюється (реплицируется). Завдяки цьому у всіх дочірніх клітин також маються ядра.
Будова ядра різна в стані поділу і між поділами. Останнє називається часто спочиваючим станом, хоча це по суті невірно, так як в цей період в ядрі найбільш активно відбуваються процеси обміну речовин. Краще називати таке "спочиваюче" ядро інтеркінетіческім, тобто знаходяться в проміжках між поділами. У такому стані ядра найчастіше і доводиться спостерігати. В живих, абсолютно непошкоджених клітинах вищих рослин ядра нерідко мало відрізняються по світлозаломлення від навколишнього цитоплазми, так що при спостереженні слабо помітні в вигляді бульбашки зі світлим контуром. У такому бульбашці різко виступають одне або кілька ядерець у вигляді круглих, сильніше заломлюючих світло (блискучих) тілець. Після фіксації та забарвлення ядра робляться видимі значно ясніше. Ядро фіксованою клітини, що знаходиться в інтеркінетіческом стані, містить ядерний сік, або каріолімфа, в якому розташовані тонка хроматинова нитчасті-зерниста структура - хроматин і одне або кілька ядерець, особливо яскраво окрашивающихся основними барвниками завдяки високій концентрації рибонуклеопротеидов. Від цитоплазми ядро отграничено дуже тонкої цитоплазматичної мембраною (ядерної оболонкою). Електронний мікроскоп дозволяє бачити, що мембрана двуслойной і в ній є дуже тонкі пронизують її канальці. Оболонка ядра являє собою частину мембранної системи клітини. Між ядром і цитоплазмою здійснюється постійний обмін речовин. Описані структури добре видно на фіксованих і забарвлених препаратах. Наявність описаних структур підтверджують і фотографічні знімки з живих, нефіксованих і нефарбованих ядер. За хімічним складом і фізико-хімічною будовою як ядро, так і цитоплазма представляють в основному комплекс гідрофільних колоїдів, що знаходяться в більшій чи меншій мірі набухання. З них головне значення мають білки, і особливо складні білки, звані протеидами, що представляють з'єднання власне білка (протеїну) з небілковим компонентом, здебільшого кислотного характеру. З них особливо важливе значення мають нуклеопротеїди - сполуки білка з нуклеїновими кислотами. Відомі два типи нуклеїнових кислот: що містяться в ядрі дезоксирибонуклеїнової (тімонуклеіновие), скорочено звані ДНК, і що містяться в ядерцях і цитоплазмі рибонуклеїнові кислоти, що позначаються РНК. Вони зумовлюють окрашиваемость ядра основними фарбами, тому що лежать в основі хроматіна2. ДНК вельми точно виявляються кольоровий реакцією Фельгена3. РНК цієї реакції не дають. ДНК входять до складу клітинного ядра у вигляді нуклеопротеидов, а почасти, мабуть, і у вільному стані. Нуклеїнові кислоти являють собою дуже довгі ланцюжки, ланки яких - нуклеотиди - складаються з пятиуглеродного цукру (рибози C5H10O5 або дезоксирибози C5H10O4), фосфорної кислоти і одного з азотистих основ. Макромолекула нуклеїнової кислоти складається з двох таких ланцюгів, обвивающих одна іншу і створюючих таким чином подвійну спіраль. Кожна стрічка представляє собою основну ланцюг. Горизонтальні відрізки відповідають парам підстав, приєднаних по одному до кожної ланцюга. Ядро здійснює дві групи загальних функцій: одну, пов'язану власне з зберіганням генетичної інформації, іншу - з її реалізацією, із забезпеченням синтезу білка.
В першу групу входять процеси, пов'язані з підтриманням спадкової інформації у вигляді незмінної структури ДНК. Ці процеси пов'язані з наявністю так званих репараційних ферментів, які ліквідують спонтанні пошкодження молекули ДНК (розрив однієї з ланцюгів ДНК, частина радіаційних ушкоджень), що зберігає будова молекул ДНК практично незмінним в ряду поколінь клітин або організмів. Далі, в ядрі відбувається відтворення або редуплікація молекул ДНК, що дає можливість двом клітинам отримати абсолютно однакові і в якісному і в кількісному сенсі обсяги генетичної інформації. В ядрах відбуваються процеси зміни і рекомбінації генетичного матеріалу, що спостерігається під час мейозу (кросинговер). Нарешті, ядра безпосередньо беруть участь у процесах розподілу молекул ДНК при діленні клітин. Отже, в ядрі клітин укладені хромосоми, які містять ДНК - сховище спадкової інформації. Цим визначається провідна роль клітинного ядра в спадковості. Дане найважливіше положення сучасної біології не просто витікає з логічних міркувань, воно доведено рядом точних дослідів. Наведемо один з них. У Середземному морі мешкає декілька видів одноклітинних зелених водоростей - Ацетабулярія. Вони складаються з тонких стеблинок, на верхніх кінцях яких розташовуються капелюшки. За формою капелюшків розрізняють види Ацетабулярія. В нижньому кінці стеблинки ацетабулярії знаходиться ядро. У ацетабулярії одного виду штучно видалили капелюшок і ядро, а до стеблинки підсадили ядро, витягнуте у ацетабулярії іншого виду. Що ж виявилося? Через деякий час на водорості з підсадженим ядром утворилася капелюшок, характерна для того виду, якому належало пересаджене ядро. Хоча ядру належить провідна роль в явищах спадковості, з цього, однак, не випливає, що тільки ядро відповідально за передачу всіх властивостей з покоління в покоління. В цитоплазмі також існують органели (хлоропласти і мітохондрії), що містять ДНК і здатні передавати спадкову інформацію. Таким чином, саме в ядрі кожної клітини міститься основна спадкова інформація, необхідна для розвитку цілого організму з усім розмаїттям його властивостей і ознак. Саме ядро відіграє центральну роль в явищах спадковості. Інший групою клітинних процесів, що забезпечуються активністю ядра, є створення власне апарату білкового синтезу. Це не тільки синтез, транскрипція на молекулах ДНК різних інформаційних РНК і Хвороби. В ядрі еукаріотів відбувається також утворення субоедениц рибосом шляхом комплексування синтезованих в полісом рибосомних РНК з рибосомні білками, які синтезуються в цитоплазмі і переносяться в ядро.Таким чином, ядро являє собою не тільки вмістилище генетичного матеріалу, а й місце, де цей матеріал функціонує і відтворюється. Тому випадання лив порушення будь-якої з перерахованих вище функцій згубно для клітини в цілому. Так порушення репараційних процесів буде приводити до зміни первинної структури ДНК і автоматично до зміни структури білків, що неодмінно позначиться на їх специфічної активності, яка може просто зникнути або змінитися так, що не забезпечуватиме клітинні функції, в результаті чого клітина гине. Порушення редуплікаціі ДНК призведуть до зупинки розмноження клітин або до появи клітин з неповноцінним набором генетичної інформації, що також згубно для клітин. До такого ж результату приведе порушення процесів розподілу генетичного матеріалу (молекул ДНК) при діленні клітин. Випадання в результаті ураження ядра або в разі порушень будь-яких регуляторних процесів синтезу будь-якої форми РНК автоматично призведе до зупинки синтезу білка в клітині або до грубих його порушень.
Значення ядра як сховища генетичного матеріалу і його головна роль у визначенні фенотипічних ознак були встановлені давно. Німецький біолог Хаммерлінг одним з перших продемонстрував найважливішу роль ядра. Він вибрав як об'єкта своїх експериментів надзвичайно велику одноклітинних (або неклеточную) морську водорість Acetabularia. Існує два близько споріднених виду A. medierranea і A. crenulata, що розрізняються тільки за формою УшляпкіФ. У ряді експериментів, у тому числі таких, в яких УшляпкуФ відокремлювали від нижньої частини УстебелькаФ (де знаходиться ядро), Хаммерлінг показав, що для нормального розвитку капелюшки необхідно ядро. У подальших експериментах, в яких з'єднували нижню частину, що містить ядро одного виду з позбавленим ядра стебельком іншого виду, у таких химер завжди розвивалася капелюшок, типова для того виду, якому належить ядро. При оцінці цієї моделі ядерного контролю слід, однак, враховувати примітивність організму, використаного в якості об'єкта. Метод пересадок був застосований пізніше в експериментах, проведених в 1952 р двома американськими дослідниками, Бріггсом і Кінгом, з клітинами жаби Rana pipenis. Ці автори видаляли з незапліднених яйцеклітин ядра і замінювали їх ядрами з клітин пізньої бластули, вже виявляли ознаки диференціювання. У багатьох випадках з яєць реципієнтів розвивалися нормальні дорослі жаби.7. Ендоплазматична сітка. Хімічний склад.Ендоплазматична сітка (від грец. endos — внутрішній), або ендоплазматичний ретикулум (від лат. reticulum — сітка) — це мембранна органела, яка ділить цитоплазму на компартменти (від англ. сompartment — відділ, відсік). Ендоплазматична сітка — це порожниста система у вигляді замкненої сукупності канальців і цистерн, утворених суцільною безперервною мембраною і заповнена матриксом (рис. 2.21). Матрикс (від лат. matrix — мати) — це пухкий матеріал помірної щільності (продукт синтезу). Ендоплазматична сітка відкривається в перинуклеарний простір — простір між двома мембранами каріолеми. Це — синтетичний і частково транспортний апарат цитоплазми, що забезпечує виконання клітиною складних функцій.Біохімічний склад мембран ендоплазматичної сітки, виявлений шляхом диференціального центрифугування, показав, що сітка містить білки і ліпіди, серед яких багато фосфоліпідів, а також ферменти: аденозинтрифосфатазу, ферменти синтезу мембранних ліпідів. При виготовленні мікроскопічних препаратів цитоплазма клітин з добре розвиненою гранулярною ендоплазматичною сіткою фарбується основними барвниками. У класичній гістології базофільна субстанція (тигроїд, або субстанція Нісля), описана в нейронах, є скупченням угрупувань гранулярної ендоплазматичної сітки.Розрізняють два види ендоплазматичної сітки: гладку (агранулярну, аЕС), представлену трубочками, що анастомозують між собою, і шорстку (гранулярну, грЕС), побудовану з цистерн, також з’єднаних між собою і вкритих полісомами. Деякі автори виділяють ще перехідну, або транзиторну (тЕС) ендоплазматичну сітку, яка знаходиться в ділянці переходу однієї різновидності ЕС в іншу.Гранулярна ЕС присутня у всіх клітинах (за винятком сперматозоїдів), однак ступінь її розвитку різний і залежить від спеціалізації клітин. Так, сильно розвинену грЕС мають епітеліальні залозисті клітини (підшлункової залози, які виробляють травні ферменти, печінкові — синтезують альбуміни сироватки крові), фібробласти (клітини сполучної тканини, які продукують білок калоген), плазматичні клітини (продукують імуноглобуліни).Агранулярна ЕС переважає в клітинах надниркових залоз (здійснює синтез стероїдних гормонів), в м’язових клітинах (бере участь в обміні кальцію), в клітинах фундальних залоз шлунка (участь у виділенні Сl–).Функції ендоплазматичної сітки. Гладка (агранулярна) і шорстка (гранулярна) ендоплазматичні сітки своїми мембранами і вмістом (матриксом) виконують чотири спільні функції:(1) розмежувальну, структуруючу, яка забезпечує упорядкований розподіл цитоплазми, не допускає її змішування та потрапляння в органелу випадкових речовин;(2) трансмембранний транспорт, завдяки якому здійснюється перенесення через стінку мембрани необхідних речовин;(3) синтез мембранних ліпідів ферментами, які містяться в самій мембрані і забезпечують репродукцію ендоплазматичної сітки;(4) вважають, що різниця потенціалів, яка виникає між двома поверхнями мембран органели можливо забезпечує проведення імпульсів збудження.Крім того, кожна з різновидностей сітки виконує свої спеціальні функції.Агранулярна ендоплазматична сітка, крім названих функцій, спільних для обох видів ЕС, виконує ще й властиві лише їй функції:(1) Депо кальцію. У багатьох клітинах (скелетних і серцевих м’язах, нейронах, яйцеклітині) існують механізми, які забезпечують зміну концентрації Са2+, такі як Са2+-помпа, Са2+-канал, Са2+-зв’язувальний білок. У поперечнопосмугованій м’язовій тканині існує спеціалізована гладка ендоплазматична сітка, названа саркоплазматичним ретикулумом, яка є резервуаром Са2+, а її мембрани містять потужні кальцієві помпи, які в соті частки секунди можуть викидати велику кількість кальцію в цитоплазму або транспортувати його у порожнини каналів ретикулума (детальніше описана в курсі гістології в розділі про поперечносмугасту м’язову тканину).(2) Синтез ліпідів, речовин типу холестерину і стероїдних гормонів. Синтез стероїдних гормонів відбувається головним чином в ендокринних клітинах гонад і надниркових залоз, а також в клітинах печінки, нирок. У клітинах кишечника синтезуються ліпіди, які у вигляді хіломікронів виводяться в лімфу, а потім у кров.(3) Детоксикуюча функція — знешкодження екзогенних і ендогенних токсинів. Наприклад, в печінкових клітинах (гепатоцитах) є ферменти оксидази, які руйнують фенобарбітал.(4) Ферменти органели беруть участь у синтезі глікогену (у печінкових клітинах).(5) Дещо видозмінена гладка ендоплазматична сітка добре розвинена в парієтальних клітинах фундальних залоз шлунка. Там вона бере участь у механізмі, який забезпечує підтримання концентрації іонів хлору, необхідних для вироблення соляної кислоти як складової шлункового соку.Гранулярна ендоплазматична сітка, крім названих спільних, виконує ще спеціальні функції.(1) Синтез білків на грЕС має деякі особливості (рис. 2.22). Починається він на вільних полісомах, які в дальшому зв’язуються з мембранами ЕС. На першому етапі взаємодії іРНК з рибосомами відбувається утворення особливого сигнального пептида, який контактує з сигнал-розпізнавальною частиною (СРЧ). Комплекс СРЧ-рибосома зв’язується з рецептором на мембрані ЕС, який називається причальним білком. Крім того на мембрані грЕС є інтегральні рецепторні білки рибофорини (зв’язані з порою), що забезпечують прикріплення великої субодиниці, а тим самим і всієї рРНК. Після цього сигнал-розпізнавальна частина відділяється від рибосоми і на рибосомі починається синтез білка. Під дією сигнальної пептидази від’єднується сигнальний пептид і синтез білка продовжується, а білок через пору в мембрані ЕС виділяється в її просвіт. Після закінчення синтезу субодиниці рибосом дисоціюють, звільнюються від іРНК і можуть знов включатися в процес синтезу.грЕС синтезує всі білки клітинних мембран (за винятком деяких гідрофобних білків внутрішніх мембран мітохондрій і хлоропластів), специфічні білки, розміщені у внутрішній фазі мембранних органел, а також секреторні білки, які транспортуються клітиною у внутрішню обмежену мембраною фазу цитоплазми і поступають у позаклітинний простір.(2) Посттрансляційна модифікація білків: гідроксилування, сульфатування, фосфорилювання. Важливим посттрансляційним процесом є глікозилування — приєднання до білків олігосахаридів з утворенням глікопротеїдів, яке забезпечується зв’язаним з мембраною ферментом глікозилтрансферазою. Глікозилування відбувається перед секрецією або транспортом речовин до деяких ділянок у клітині (комплексу Гольджі, лізосом або плазмолеми).(3) Транспорт речовин по внутрішньомембранній фазі сітки. Синтезовані білки просвітами ЕС переміщаються до комплексу Гольджі, який бере участь у виведенні речовин з клітини.Згідно з сигнальною гіпотезою існує ряд механізмів, взаємодія яких забезпечує надходження синтезованих секреторних, мембранних і лізосомних білків в просвіт грЕС. Зокрема, в мембрані гранулярної ЕС є інтегральні рецепторні білки рибофорини, що забезпечують прикріплення більших субодиниць рибосом. Вони формують у мембрані гідрофобні канали, які служать для проникнення синтезованого білкового ланцюжка в просвіт гранулярної ЕС. На більшій субодиниці рибосом є сигнал-розпізнавальний білок. Взаємодія сигнал-розпізнавальної частини на рибосомі зі своїм рецептором на мембрані ЕС забезпечує потрапляння синтезованого білка через мембранний канал у просвіт грЕС.(4) Гранулярна ендоплазматична сітка бере участь в утворенні комплексу Гольджі.Утворення ендоплазматичної сітки. Ліпідний компонент мембран органели синтезується ферментами, розташованими в самій сітці, білковий — поступає з рибосом, що містяться на її мембранах. Гладка ендоплазматична сітка не має власних факторів для синтезу білка, тому вважається, що ця органела утворюється з гранулярної ендоплазматичної сітки, яка, втрачаючи рибосоми, переходить в агранулярну.8. Апарат Гольжді.Опис структури апарату Гольджі тісно пов'язане з описом егоосновних біохімічних функцій, оскільки підрозділ етогоклеточного компартмента на відділи виробляється переважно на основі локалізації ферментів, розташованих в тому чи іншому відділі. Найчастіше в апараті Гольджі виділяють чотири основні відділи: цис- Гольджи, медіа-Гольджі, транс-Гольджі і транс-Гольджі мережу (TGN) Крім того до апарату Гольджі іноді відносять так називаемийпромежуточний компартмент, що представляє собою скупчення мембранних бульбашок між ендоплазматичним ретикулумом і цис-Гольджі. Апарат Гольджі є дуже полиморфной органел; в клітинах різних типів і навіть на різних стадіях розвитку однієї і тієї ж клітини він може виглядати по-різному. Основні його характеристики такі:
1) наявність стопки з декількох (зазвичай 3-8) сплощені цистерн, більш-менш щільно прилеглих один до одного. Така стопка завжди буває оточена деяким (іноді дуже значним) кількістю мембранних бульбашок. В тварин клітинах частіше можна зустріти одну стопку, в той час як в рослинних клітинах їх зазвичай буває декілька; кожну з них в такому випадку називають діктіосоми. Окремі діктіосоми можуть бути пов'язані між собою системою вакуолей, утворюючи тривимірну мережу;
2) композиційна гетерогенність, що виражається в тому, що постійні (resident) ферменти неоднорідне розподілені по органелле;
3) полярність, тобто наявність цис-боку, зверненої до ендоплазматичнийретикулум і ядру, і транс-боку, зверненої до поверхні клітини (це особливо характерно для секретуючих клітин);
4) асоціація з микротрубочками і областю центриоли. Руйнування микротрубочек деполімеризує агентами призводить до фрагментації апарату Гольджі, проте його функції при цьому істотно не зачіпаються. Аналогічна фрагментація спостерігається і в природних умовах, під час мітозу. Після відновлення системи мікротрубочок розкидані по клітці елементи апарату Гольджі збираються (по мікротру-бочкам) в область центриоли, і реконструюється нормальний комплекс Гольджі.
Апарат Гольджі (комплекс Гольджі) - мембранна структура еукаріотичної клітини, в основному призначена для виведення речовин, синтезованих в ЕПР. Комплекс Гольджі був названий так на честь італійського вченого Камілло Гольджі, вперше виявив його в 1898 році. Комплекс Гольджі являє собою стопку дископодібних мембранних мішечків (цистерн), кілька розширених ближче до країв і пов'язану з ними систему пухирців Гольджі. У рослинних клітинах виявляється ряд окремих стопок (діктіосоми), в тварин клітинах часто міститься одна велика або кілька з'єднаних трубками стопок. В цистернах Апарату Гольджі дозрівають білки призначені для секреції, трансмембранні білки плазматичної мембрани, білки лізосом і т.д. Що достигають білки послідовно переміщаються по цистерн органели, в яких відбувається їх остаточне згортання, а також модифікації - гликозилирование і фосфорилирование.
Апарат Гольджі ассіметрічен - цистерни розташовані ближче до ядра клітини (цис-Гольджі) містять найменш зрілі білки, до цих цистерн безперервно приєднуються мембранні пухирці - везикули, відгалужуються від шорсткогоЕПР (ЕР), на мембранах якого і відбувається синтез білків рибосомами. Різні цистерни Апарату Гольджі містять різні резидентні каталітичні ферменти і, отже, з дозріваючими білками в них послідовно відбуваються різні процеси. Зрозуміло, що такий ступінчастий процес повинен якось контролюватися. Дійсно, що дозрівають білки «маркуються» спеціальними полісахаридними залишками (переважно маннознимі), мабуть, граючими роль своєбразним «знаку якості». Не до кінця зрозуміло, яким чином дозрівають білки переміщаються по цистернах Апарату Гольджі, в той час як резидентні білки залишаються в більшій чи меншій мірі асоційовані з одного цистерною. Існують дві взаімонеісключающіе гіпотези, що пояснюють цей механізм. Згідно з першою (1), транспорт білків здійснюється за допомогою таких же механізмів везикулярного транспорту, як і шлях транспорту з ЕР, причому резидентні білки не включаються до відбруньковуватися везикулу. Згідно з другою (2), відбувається безперервне пересування (дозрівання) самих цистерн, їх збірка з бульбашок з одного кінця і розбирання з іншого кінця органели, а резидентні білки переміщаються ретроградно (в зворотному напрямку) за допомогою везикулярного транспорту. Зрештою від протилежного кінця органели (транс-Гольджі) відбруньковуються бульбашки, що містять повністю зрілі білки.
В комплексі Гольджі відбувається
О-глікозилювання, до білків приєднуються складні цукру через атом кисню.
Фосфорилування (приєднання до білків залишку ортофосфорної кислоти).
Освіта лізосом.
Освіта клітинної стінки (у рослин).
Участь в везикулярного транспорті (формування трехбелкового потоку):
дозрівання і транспорт білків плазматичної мембрани;
дозрівання і транспорт секретів;
дозрівання і транспорт ферментів лізосом.
Апарат Гольджі. Апарат Гольджі (комплекс Гольджі) - це спеціалізована частина ЕПР, що складається з зібраних в стопки плоских мембранних мішечків. Він бере участь в секреції клітиною білків (у ньому відбувається упаковка секретується білків у гранули) і тому особливо розвинений в клітинах, що виконують секреторну функцію. До важливих функцій апарату Гольджі відноситься також приєднання вуглеводних груп до білків і використання цих білків для побудови клітинної мембрани і мембрани лізосом. У деяких водоростей в апараті Гольджі здійснюється синтез волокон целюлози.
Гольджі апарат: функції
Функцією апарату Гольджі є транспорт і хімічна модифікація надходять в нього речовин. Вихідним субстратом для ферментів є білки, що надходять в апарат Гольджі з ендоплазматичного ретикулума. Після модифікації і концентрування, ферменти в бульбашках Гольджі переносяться до «місця призначення», наприклад до місця утворення нової нирки. Найбільш активно цей перенос здійснюється за участю цитоплазматичних мікротрубочок.
Функції апарату Гольджі дуже різноманітні. До них можна віднести:
1) сортування, накопичення і виведення секреторних продуктів;
2) завершення посттрансляционной модифікації білків (гликозилирование, сульфатами і т.д.);
3) накопичення молекул ліпідів та освіта ліпопротеїдів;
4) утворення лізосом;
5) синтез полісахаридів для утворення гликопротеидов, восків, камеді, слизей, речовин матриксу клітинних стінок рослин
(гемицеллюлоза, пектини) і т.п.
6) формування клітинної пластинки після ділення ядра в рослинних клітинах;
7) участь у формуванні акросоми;
8) формування скоротних вакуолей найпростіших.
Цей список, без сумніву, не вичерпний, і подальші дослідження не тільки дозволять краще зрозуміти вже відомі функції апарату Гольджі, а й призведуть до відкриття нових. Поки самими вивченими з біохімічної точки зору залишаються функції, пов'язані з транспортом і модифікаціє новосинтезованих білків.