

фізіологія Плиска остання
.pdfтрофотропну й ерготропну дії. Окремим випадком трофотропної регуляції є участь гормонів у регуляції процесів росту й розвитку організму з метою забезпечення його лінійного росту та процесів диференціювання.
Однією з відмітних характеристик гормонів є їхня дистантна дія. Потрапляючи в кров, гормони, як правило, впливають не на найближчі, а на віддалені органи. Ендокринні клітини, які виділяють гормони, відносно регулювальних органів відіграють роль центрального (дистантного) механізму регуляції. Тому гуморальну регуляцію, яка забезпечується гормонами, часто називають централь-
ною, або дистантною регуляцією.
На відміну від гормонів, гормоноїди і метаболіти, виділяючись у кров, впливають лише на найближчі органи. Тому ці речовини є чинниками місцевої дії. Відповідно гуморальну регуляцію, яку во-
ни забезпечують, називають місцевою гуморальною регуляцією.
Різні гуморальні фактори можуть спільно впливати на одні й ті самі органи і клітини організму. До того ж їхні впливи можуть бути як синергічними, так і антагоністичними.
Дія гуморальних факторів на клітини організму завжди здійснюється за участю «молекулярних циторецепторів»: цитоплазматичних, мембранних або ядерних.
Молекулярний циторецептор (рецетор) — це група молекул ци-
топлазми або мембрани клітини, які мають вибіркову чутливість до певної хімічної речовини (гуморального фактора, медіатора). Для нього характерні просторова й структурна специфічність до агоніста, обмежена кількість місць зв'язування, висока спорідненість, оборотне зв'язування. Структура рецептора складається з білкової макромолекули та багатокомпонентного небілкового комплексу з можливістю динамічного комплексоутворення. Кількість і спорідненість рецепторів до гормонів регулюються короткочасними й довготривалими механізмами. Це підтверджує, що всі компоненти плазматичної мембрани перебувають у динамічній рівновазі. Вважають, що тривала зайнятість більшості рецепторів певними гормонами викликає їх ендоцитоз і руйнування. Це захищає клітину від надмірної стимуляції. Максимальний ефект спостерігається при зайнятості 5-10 % загальної кількості дільниць зв'язування. Інші є запасними (резервними) або не мають зв'язку з комплексом ефекторних молекул, що забезпечують виявлення біологічного ефекту. Розрізняють іонотропні (І класу) та метаболотропні (II класу) рецептори. Активація перших — КА, ацетилхоліном спричиняє розвиток надшвидких іонних струмів; других — активує ефекторні клітини через G-білок і реакції розвиваються більш повільно — гіпоталамогіпофізарні гормони.
133

Розрізняють два механізми дії хімічних речовин на клітини організму:
1. Дія хімічних речовин на клітини через їхні цитоплазматичні рецептори (рис. 40).
У цьому разі БАР проникає крізь мембрану клітини і з'єднується з її цитоплазматичними рецепторами (складається із субодиниць А і В), вибірково чутливими до цього фактора. Утворений комплекс трансформується і транслокується (переміщується) в ядро,
зв'язується з хроматиновим фактором і субодиниця А впливає на генетичний апарат (ДНК) зі зміною швидкості транскрипції та кількості інформаційної (матричної), транспортної і рибосомної рибонуклеїнової кислоти (РНК). Матрична, або інформаційна, РНК переносить інформацію з ядра в цитоплазму й безпосередньо бере участь у збиранні молекули поліпептиду рибосомами на шершавому ЕР, транспортна — доставляє амінокислоту до рибосоми, рибосомна — взаємодіє з попередніми двома вході збиранняполіпептиду і в комплексі з білками (в тому числі й ферментами) утворює рибосому. Як наслідок, змінюється інтенсивність утворення клітиноюпевнихбілкових речовин: білків-ферментів, транспортнихсистем, структурних білків тощо. Це виявляється в суворо визначених біологічних ефектах, які є специфічним результатом дії даної хімічної речовини на клітину. Ці ефекти полягають у підвищенні проникності клітинної мембрани для певних іонів, посиленні енергозабезпечення клітини, зміні параметрів збудження або параметрів специфічної функції даної клітини. Субодиниця В зв'язується з негістоновими білками. Можливо, стероїди спочатку зв'язуються зі специфічними білками мембрани клітини, які транспортують їх до цитоплазматичногорецептораабоповзнього— дорецепторівядра.
Тироксин може прямо проникати в ядро й там зв'язуватися з рецептором. Надалі так само.
Такий механізм дії на клітину хімічних речовин властивий речовинам з невеликою молекулярною масою і здатним проникати через клітинну мембрану (стероїдні гормони, гормони щитоподібної залози та інші небілкові гормони). Стероїдні гормони розчиняються в ліпідному бішарі. Сучасна теорія доводить, що у плазматичній мембрані наявні специфічні рецептори для естрогенів і глюкокортикоїдів, які транспортують відповідні гормони.
134

Ранні й пізні ефекти цих гормонів пов'язані з теоріями «доміно» і «безперервної дії». За першою — комплекс посилює синтез кількох типів мРНК, які транслюються з утворенням нових білків. Ця реакція викликає другу хвилю вибіркового синтезу білка. За другою теорією, ініціація пізніх ефектів потребує не тільки синтезу РНК і накопичення специфічних білків на ранньому етапі реакції (протягом години), а й безперервної присутності гормон-рецепторного комплексу в ядрі протягом критичного періоду, який виходить за межі ранніх реакцій.
2. Дія хімічних речовин на клітини через їхні мембранні циторецептори і внутрішньоклітинні посередники.
Внутрішньоклітинні посередники (месенджери)- це певні хіміч-
ні сполуки, які посилено утворюються в клітинах при дії на їх мембранні рецептори різних гуморальних факторів або медіаторів і які реалізують усередині клітин впливи цих речовин.
Залежно від внутрішньоклітинного посередника розрізняють декілька підвидів такої дії хімічних речовин:
1) Гормони (хімічні, або первинні посередники), або хімічні речовини, які діють на клітину через Ац систему (рис. 41). До них відносять КА, що зв'язуються з р-адренорецепторами, вазопресин, що зв'язується з У2-рецепторами, глюкагон, паратирин, кальцитонін, секретин, гістамін, що зв'язуються з Н2-рецепторами.
Розглянемо цей механізм дії на прикладі адреналіну. Відбувається такийланцюгпослідовнихпроцесів, щовипливаютьодинзодного:
а) Взаємодія адреналіну з р-адренорецептором клітинної мембранийактивація; черезр2-адренорецептори— гальмування.
б) Активація Ац — ферменту, розташованого на внутрішній поверхні клітинної мембрани.
Здійснюєтьсячерезбілковийкомпонентмембрани— G-білок(N-білок). Цетрансдуктор(тример), якийскладаєтьсяза-, р- іу-субодиниць. а-Суб- одиницямаєГТФ-азну активність. Активація
G-білка й передача сигналунакаталітичну субодиницюАцсупро-
воджуютьсягідролізом |
4ТФ- -*-uA |
ГТФ до ГДФ + Фн. |
♦ |
Взаємодія3 гормонами |
nf фосфорияюватя Певнийбіологічний |
викликає ДИСОЦіаціЮ |
""певного білка ~т'ефект клітини |
перших двох одиниць |
|
і дисоціацію комп- |
Рис. 41. Взаємодія гормонів з мембранним ци- |
лексу G-білок — ГДФ |
торецептором (аденілатциклазний механізм |
та взаємодію G-білка |
взаємодії): ПК — протеїнкіназа |
|
135 |
з ГТФ. Утворений комплекс G-білок — ГТФ приєднує каталітичну субодиницю, утворюючи активну Ац. Існує різних 16 — а-, 4 — р- і7 — у-субоди- ниць. Вони дають 1000 комбінацій. Крім того, розрізняють Gs (стимулювальні — активують Ац), G, (інгібують Ац), Gq (активують фосфоліпазу С) і Gt види білка.
в) Посилене утворення в клітинах з АТФ-Mg2* циклічного адено- зин-3'5'-монофосфату (цАМФ), який є найбільш вивченим внутрішньоклітинним (вторинним) посередником.
Водночас комплекс G-білок — ГТФ спричиняє дисоціацію гормонрецепторного комплексу шляхомзниженняспорідненості рецепторадогормонів.
Циклічний АМФ посилено утворюється вклітинахпридіїнаїхні мембранні рецептори багатьох гормоніві медіаторівта, як приклад, КА(адреналіну й норадреналіну). Кожна з цих речовин сприймається «своїми» мембранними рецепторами, тобторецепторами, вибірково чутливими тільки до даної речовини. Зв'язуваня ліганду з Gsa — сильно активує, Gj(x — слабо гальмує Ац, і відрізняються вони a-субодиницею, Gc також є «трансдуктором» — перетворювального елемента в процесі активації фосфодіестерази ГМФ під впливом світла.
Аденілатциклаза |
Фосфодіестераза |
I |
I |
АТФ -► цАМФ (3'5' - АМФ) + ФФ„; |
цАМФ -► 5' - АМФ |
Останню реакцію блокує роліпрем (селективний інгібітор фосфодіестерази); сама фосфодіестераза гальмує дію метилксантинів (кофеїну, теофіліну).
г) Активація внутрішньоклітинних цАМФ-залежних потеїнкіназ — ферментів, які фосфорилюють різні білки клітини. Фосфат приєднується до білків кіназами та відщеплюється фосфатазами.
д) Фосфорилювання білків зовнішньої мембрани клітини, білків трансмембранних переносників, білків ферментів тощо. Так, у клітинах міокарда Ац система активує Л-кінази, які фосфорилюють білки кальцієвих каналів мембрани кардіоміоцитів. Це призводить до збільшення їхньої провідності. У результаті збільшується вхід Са2+ з позаклітинного простору в цитоплазму кардіоміоцитів під час їх збудження. Вхід Са2+ сам по собі активує певні внутрішньоклітинні білки, викликаючи специфічні ефекти. Таким чином, у цьому разі з'являється ще один внутрішньоклітинний (третинний) посередник— Са2+.
Фосфат приєднується до білків кіназами і відщеплюється фосфатазами.
136
2) Гормони, які діють на клітини через Са2+-поліфосфоінозитоль- ну систему. Це — КА, що зв'язується з а-АР; вазопресин, що зв'я- зується з V,-рецепторами; окситоцин, гастрин, панкреозимін, гістамін, серотонін, що зв'язуються з Н,-рецепторами.
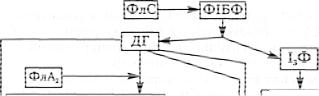
У цьому разі гормон взаємодіє з відповідними мембранними рецепторами клітини. Це активує мембранну фосфоліпазу С (фермент). Остання спричиняє гідроліз мембранного фосфоліпіду — поліфосфоінозитиду (фосфатидилінозитолу) з утворенням двох внутрішньоклітинних посередників (каскад реакцій зображено нарис. 42).
Діацилгліцерол (ДГ) — ліпофільний і залишається біля мембрани, викликаючи вхід Са2+ через низькопорогові Са2+-канали. І3Т — водорозчинний, дифундує до СР та спричиняє вихід Са2+ з внутрішньоклітинних депо. Гуанікатциклаза (Гц) є зв'язана з мембраною і водорозчинна. Кальмодулін може зв'язувати чотири іони кальцію.
Простаноїди — це похідні простанової кислоти: простагландини (А, В, D, E, F, G, Н, І), простацикліни та тромбоксан А2.
При цьому спостерігаються «партнерство», «подвійний» (страхувальний) контроль, комплексування, послідовність та антагонізм дії посередників. Простагландини, тромбоксани та простацикліни утворюються з ендоперекисів арахідонової кислоти й мають короткий період напіврозпаду. Ейкозаноїди (ауто- і паракринні фактори) посилюють або послаблюють діюгормонівтаіншихагоністів, щовикликаютьїхсинтезісекреціютаєтакож посередниками. Простагландини — біологічно активні речовини, похідні а- та у-лінолевої кислоти, синтезовані в клітинах, які діють через Ац систему. Локалізуютьсявонивосновному йліпідномушарімембрани. Вони швидко руйнуються в легенях, нирках, печінці. Метта лейенкефаліни локалізуються в тканинах вісцеральних органів. Так, лейенкефалін синтезується в ендокринних клітинах ШКТ, мозковому шарі наднирників, гангліях симпатичної та метасимпатичної нервової системи. Рецептори до нихподіляютьсянамю-, дельта-, капа-, сігма-, епсілон-підгрупийлокалізо- вані як на зовнішній, так і на внутрішній поверхні плазматичної мембрани. Механізм дії пов'язаний з гальмуванням входу Са2+ в пресинаптичну мембрану термінальних нейронів зі зниженням виділення ацетилхоліну, норадреналіну, субстанції Р та гальмуванням збудження в синапсах. Даларгін (аналог лейенкефаліну) застосовують в анестезіології та гастроентерології.
Блокатори ліпооксигенази — вітамін Е, кверцетин (карвітин), циклооксигенази — індометацин, аспірин. Блокування одного із шляхів значно модулюєфункціїорганізму.
Простациклін бере участь у процесах агрегації та дезагрегації тромбоцитів, підвищення [Са2+]„ активує кальцієві насоси та канали, які нормалізують внутрішньоклітинну концентрацію Са2+, викачуючи його в міжклітинний простір і СР з відновленням функціонального спокою. Кальмодулін також діє на Ац, Гц, фосфодіестеразу, фосфорилкіназу,
137
ГідролізДГзутворенням арахідонової кіслоти
циклооксигеназа лшооксгеназа
ПР-Прс (А, В, Е, F, І) Лкт(А, В, С, ПстІ, ТрAj
|Ейкозаноїди|
АктиваціямембранногоферментуГц
Накопичення в цитоплазмі внутрішньоклітинного посередника цГМФ
(ГТФ -> цГМФ)
Активація ПКпевного |
АктиваціяПк певного |
виду (У-кінази) |
виду (С-кінази) |
Фосфорилювання |
І |
Фосфорилювання |
|
певних білків |
певнихбілків клітини |
КЛІТИНИ |
(заучастюСа2+) |
Дифузія І3Фвцитоплазму істимуляціявиходузЕРСа2+ як внутрішньоклітинного посередника
ПідвищеннявцитоплазміСа2+
З'єднанняСа2+ зKM ііншими кальційсенсорними білками, щоведедоутворення активнихкомплексів
Безпосереднє Са2+КМ- з'єднанняСа2+ неактивний з певними
білкамиклітини
Активація ПК Са2++КМ- різних видів активний
К-Ф Фосфорилюванняпевнихбілків
ї
Зміна активності різнихбілків клітини внаслідок зміни конфігурації їхмолекул
| Певні біологічні ефекти, властиві даному гормону |
Рис. 42. Схема певних біологічних ефектів, властивих даному гормону:
ФлС — фосфоліпаза С; ФІБФ — фосфоінозитолбіфосфат, KM — кальмодулін; К-Ф — кіназа-фософорилаза; ПР — простаноїди: Пре — простагландини, Пст — простациклін І; Тр А2 — тромбоксан А2; Лкт — лейкотрієни (за Тейлором)
міозинкіназу, фосфоліпазу А2, Са2+- і Mg2+-AT<D-a3y; стимулює звільнення нейротрансмітерів, фосфорилювання білків мембран. Кальмодулін — це аналог м'язового білка тропоніну С. При цьому зміну активності ферменту, що діє в ключовій точці каскаду реакцій, визначають: а) загальна кількість молекул ферменту, яка є постійною, але активність їх зростає чи зменшується внаслідок взаємодії з алостеричним або ковалентним модулятором
138
(PO4Z ), що призводить до змін спорідненості ферменту до реактантів; б) загальнакількістьмолекулферменту, якомуможепередуватитасамамодуляторнареакція, щоівпершому разі(довготривалаабохронічнарегуляція).
Необхідно зазначити, що Са2+ активує кальмодулін. Останній активує фосфодіестеразу. Глюкокортикоїди (стероїди) гальмують фосфоліпазу А2 — фермент, який мобілізує арахідонову кислоту. Це викликає протизапальну дію.
Запалення— комплексмісцевихреакційнапошкодженнятканин. Синтез простагландинів, які медіюють запалення, блокується аспірином. Повільного реагування субстанція — метаболіт арахідонової кислоти —
у400 разівсильніше дієнамускулатуру трахеїтабронхів, ніжгістамін.
3)Гормони, для яких внутрішньоклітинний посередник ще не виявлений, — соматотропний гормон і соматомедіни, інсулін.
Гуморальна метаболічна регуляція виникла на перших етапах розвитку життя. Потім з'явилася місцева гормональна регуляція. І лише значно пізніше у тварин з досить розвинутоюнервовоюсистемою формується дистантна гормональна регуляція.
Однак усі механізми гуморальної регуляції функціонують у тісному взаємозв'язку з нервовими центрами, являючи собою в сукуп- ностієдинийнервово-гуморальнийапаратрегуляціїорганізму. Причому в цьому апараті провідна роль належить нервовим механізмам.
У разі гуморальної регуляції нервові центри можуть впливати на ендокринні залози безпосередньо (через нервові волокна) або через інші ендокринні залози, простіше за все через гіпофіз.
Як результат, контури гуморальної регуляції не є самостійними, а завжди входять до складу контурів вищого порядку — контурів нервово-гуморальної регуляції. У кожному з таких контурів роль керуючого пристрою відіграє нервовий центр разом з ендокринною залозою (або кількома ендокринними залозами) (рис. 43).
Механізми місцевої гуморальної регуляції ворганізмі єбільшсамостійними, ніж механізми гормональної, тобто центральної гуморальної регуляції. Контур місцевої гуморальної регуляції далеко не завжди формує сумісно з нервовими механізмами єдиний контур — контур нервово-гуморальної регуляції.
Керуючийпристрій: нервовийцентр— ендокринназалоза
Пристрійстеження |
Виконавчімеханізми |
Пристосовнийрезультат
Рис. 43. Схеманервово-гуморального контурурегуляції
139
Частіше параметри організму, які регулюються місцевими гуморальними механізмами (контурами місцевої гуморальної регуляції), паралельно можуть регулюватись і нервовими центрами (контурами нервової або нервово-гуморальної регуляції). Якщо місцеві гуморальні механізми не можуть забезпечити бажаного пристосовного результату, то цей результат досягається за рахунок діяльності нервових або нервово-гуморальних механізмів. При цьому метасимпатична нервова система виконує роль колектора — переключає та трансформує імпульси, які поступають через симпатичні та парасимпатичні волокна.
Слід зазначити, що гуморальні механізми, відіграючи підпорядковану роль відносно нервових, у свою чергу можуть впливати на їх діяльність: а) гуморальні фактори можуть викликати в організмі різні ефекти (діючи на хеморецептори або безпосередньо на нервові центри); б) гуморальні фактори можуть змінювати силу рефлексів (коригуючи збудливість рефлекторних центрів або змінюючи чутливість органів ефекторів до медіатора).
Ендокринні залози та їхні гормони. Вегетативне забезпечення організму під час м'язової діяльності
■«Ніщоневиснажуєінеруйнуєлюдину, якдовготривала фізична бездіяльність. Ніякі ліки не замінятьрух, рух заміняє багато ліків».
Арістотель
Будь-яка діяльність цілісного організму забезпечується різними функціональнимисистемами, зякиху кожному конкретному випадку одна з них є головною, інші — обслуговувальними. Приміром, при м'язовій діяльності організму головною функціональною системою є система опори і рухів, роль обслуговувальних відіграють вісцеральні системи організму. Це — системиживлення, дихання, транспорту. До останньої входятьу цьому разіпідсистемикрові тасерце- во-судинна. Робота всіх зазначених систем при м'язовій діяльності організму спрямована на вегетативне забезпечення м'язової роботи, тобто на вегетативне забезпечення системи опори й рухів. Усі вегетативні зсуви при м'язовій діяльності стосуються, головним чином, процесів її енергозабезпечення. Мета — посилення м'язових процесів для роботи. Найшвидше проходить мобілізація енергетичних ресурсів самих м'язових клітин, органів депо системи живлення,
атакож систем дихання і кровообігу.
Ужиттєво важливих ситуаціях, навпаки, головною функціональною системою може виступати та або інша вісцеральна система,
140
а система опори і рухів відіграватиме роль обслуговувальної. її мета тепер полягатиме у забезпеченні такої вісцеральної системи. Наприклад, при голодуванні організму головною системою є система живлення. У цьому разі система опори і рухівзабезпечуватиме формування поведінкових рухових дій для здійснення вегетативних функцій.
Таким чином, інтеграція рухових і вегетативних функцій організму може відбуватися за двома напрямками (узгодження вісцеральних систем і системи опори і рухів): а) вегетативного забезпечення функцій системи опори і рухів при м'язовій діяльності та б) формування (запуску) рухового поведінкового акту з метою забезпечення вегетативних функцій.
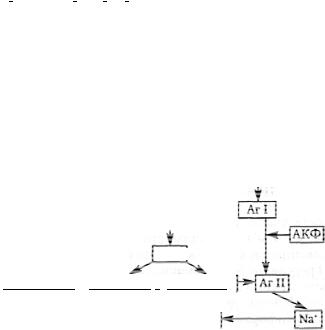
Розглянемо механізми вегетативного забезпечення системи опори і рухів, тобто забезпечення м'язової діяльності (рис. 44). Прим'я- зовій роботі також активуються внутрішні захисні реакції організму (прискорюється процес зсідання крові, стимулюється імунна відповідь організму). Мабуть, усе починається з лобної кори, яка, вважають, бере участь у формуванні мотивацій організму і, можливо, запускає його м'язову діяльність, активуючи розміщену у сусідстві сенсомоторну кору. До основних функцій лобної кори також відносять і регуляцію вегетативного забезпечення організму при м'язовій діяльності, яку здійснюють її інтегративні центри.
Свої функції лобна кора здійснює, використовуючи всебічну сенсорну інформацію, що надходитьдонеїчерезчисленні аферентні зв'язки від лімбічної та сенсомоторної ділянок кори. З ними вона межує і має двосторонні зв'язки. Відомо, що лімбічна кора (її стародавня частина) являє собою кірковий центр нюхової чутливості, прецентральна зона сенсомоторної кори є кірковим центром пропріорецептивної чутливості, а постцентральна зона сенсомоторної кори — кірковим центром шкірної чутливості.
Вегетативні центри лобної кори відносно вегетативних центрів гіпоталамуса є нервовими центрами вищого порядку. Тому лобна кора активує через лімбічні шляхи симпатичні ерготропні центри гіпоталамуса з метою попередньої підготовки організму до м'язової роботи. Імовірно це пов'язано з тим, що від сенсомторної кори починаються пірамідні і всі екстрапірамідні шляхи, що завжди стимулює м'язову діяльність організму. Проте частина цих шляхів фор-
мує ще і вегетативний пірамідний шлях, що йде до вегетативних центрів головного мозку. Це сідчить, що лобна кора може мобілізувати вегетативні ресурси ще і через сенсомоторну кору.
Гіпоталамус входитьдоскладу лімбічної системи. Останняскладається з поясної звивини, гіпокампа ігіпокампової звивини, передніх ядер таламуса, мигдалеподібних ядер. Лімбічна система тісно
141
Регуляція вегетативних функцій організму при м'язовій діяльності
|
|
|
|
Лобна кора |
|
|
|
|
|
|
|
|
|
->J Сенсомоторна кора |
||||||
|
Дихальний центр}<—| Лімбічна система |
■+J[ГГАКС |
||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Дихальнім'язиІ |
|
|
Гіпоталамус |
Тропнігормони |
|||||||||||||||
Підшлункова |
Вегетативні центри |
|
|
Соматокорти- |
Аксональний |
|||||||||||||||
|
|
залоза |
(АНС) Стовбур ГМ |
|
|
коліберин |
транспорт. АДГ |
|||||||||||||
|
|
|
|
|
|
|
|
СНС(САС) |
|
V V |
|
І |
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Юксагло- |
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Нейрогіпофіз. |
|||
Глюкагон Спинний мозок Аденогіпофіз |
||||||||||||||||||||
|
|
|
|
|
|
—г |
|
|
ж |
|
|
|
мерулярний |
АДГ |
||||||
|
|
|
|
|
|
|
|
|
|
|
апарат |
(Вазопресин) |
||||||||
|
|
Мозкова речовина |
Сомато- |
|
|
|
|
|
||||||||||||
|
|
|
|
наднирників |
тропний |
|
|
|
Ренін |
|||||||||||
|
|
|
|
|
|
|
ї |
|
гормон |
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|Катехоламіни| |
|
|
|
|
|
|
|
|
Кора |
|
|||||
|
|
|
|
|
|
|
|
|
ІПечінка] |
|
|
|||||||||
|
|
Сенсорні системи |
наднирників |
|
||||||||||||||||
|
|
|
|
|
(рецептори, |
|
|
|
|
|
|
|
|
Зони |
|
|
||||
|
|
|
нервові центри) |
|
|
|
|
|
|
|
|
|
|
|||||||
|Соматомедіни| | Жмутикова | |Клубочкова
Кортизол Ас
Вегетативні функції організму (неспецифічнізахисні-пристосувальні реакції, які забезпечують адаптацію організму до стресової ситуації)
Рухові функціїорганізму
Рис. 44. Вегетативне забезпечення рухових реакцій організму
пов'язана з регуляцією вегетативних функцій організму, й її раніше називали вісцеральним мозком, а гіпоталамус характеризували як вищий центр вегетативної регуляції вісцеральних функцій організму. Однак, крім цих функцій, лімбічна кора виконує й ряд інших, які відносяться до психічних. Зокрема, вона бере участь в організації сну, пам'яті, емоцій, мотивацій.
Усі ланки лімбічної системи кільцеподібно зв'язані між собою. Тому будь-яка інформація, щонадійшла дооднієїструктури, од-
разу ж передається до інших.
Гіпоталамус у складі лімбічної кори відіграє роль її «ефекторної зони» (або «вихідних воріт»). Це обумовлено тим, що в ньому
142