

Звягина Учебное пособие
.pdfперенос протонов и электронов от восстановленных коферментов НАД-зависимых дегидрогеназ на убихинон (кофермент Q). Далее комплекс I восстанавливает промежуточный переносчик кофермент Q (КоQ), который принимает водород от комплекса I. Образовавшийся КоQН2 отдает водород на комплекс III.
Комплекс III –включает в себя цитохромы в, с1 и FeS белок. Они транспортируют только электроны от КоQН2 на цитохром с. Два же протона (Н+) комплекс III выбрасывает в межмембранное пространство вместе с еще одной парой протонов, захваченной из матрикса.
Комплекс IV (цитохромоксидаза) включает в себя цитохромы а и а3. Это конечное звено в цепи электронного транспорта, оно обеспечивает перенос электронов от цитохрома с на молекулу О2 в матриксе. Также он способен захватывать 4 протона из матрикса, два из них идет на образование воды. А другие два поступают в межмембранное пространство.
Сокращенная дыхательная цепь
Комплекс II, который в укороченном варианте цепи является начальным звеном окисления, переносит электроны и протоны от ФАДН2 на мобильный убихинон (КоQ) липидного бислоя. А первое звено (комплекс I), которое имеется в полной цепи – исключается.
Перенос электронов по дыхательной цепи осуществляется в следующей последовательности:
SH2
ФАД
SH2
ФАДН2
НАД+
|
энергия |
|
|
энергия |
|
|
энергия |
|
НАДН.Н+ |
ФМН |
КоQ |
Цит b |
Цит c1 |
Цит с |
Цит a |
Цит a3 |
О2. |
Сопряжение окисления с фосфорилированием в дыхательной цепи. H+ - АТФ - синтетаза. Дыхательный контроль. Величина
Р/О. Разобщение дыхания и фосфорилирования.
За счет энергии транспорта электронов через I, III и IV комплексы дыхательной цепи из матрикса в межмембранное пространство выкачиваются протоны, что ведет к возникновению на внутренней мем-
бране митохондрий концентрационного (разница концентрации протонов) потенциала (∆рН). Так как каждый протон несет положи-
71
тельный заряд, то появляется и разность электрических зарядов по обе стороны мембраны: отрицательный – на внутренней стороне мембраны и положительный - на внешней стороне, формируется электрический потенциал (∆ψ). Согласно хемиосмотической теории Митчела, в совокупности электрический и концентрационный гради-
енты составляют электрохимический потенциал (∆μН+), который является источником для синтеза АТФ. Электрохимический потенциал и есть промежуточная форма запасания энергии.
∆μН+ = ∆ψ + ∆рН
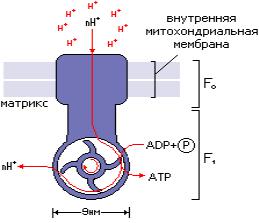
Накопившиеся в межмембранном пространстве Н+ стремятся выйти обратно в матрикс по градиенту концентраций, но митохондриальная мембрана для них непроницаема. Обратное движение протонов в матрикс возможно лишь через протонные каналы специального фермента АТФ-синтетазы (V комплекс). Этот ферментативный комплекс состоит из двух частей: Fо, которая встроена в мембрану и пронизывает ее насквозь, и F1 - напоминает шляпку гриба и обращена в матрикс митохондрии. При движении протонов через каналы F0, их энергия используется для синтеза АТФ в активном центре F1. Реакция синтеза АТФ из АДФ и Фн с использованием энергии перехода электронов и протонов по дыхательной цепи носит название окисли-
тельного фосфорилирования и описывается уравнением:
АДФ + Н3РО4= АТФ + Н2O.
Рисунок 10. Строение АТФ-синтетазы
Образовавшийся АТФ с помощью транслоказы перемещается в цитозоль; в ответ - в матрикс митохондрии поступают АДФ и фосфат.
Благодаря многоступенчатой передаче, энергия в дыхательной цепи выделяется не мгновенно, а постепенно (маленькими порциями) при каждой реакции переноса. Эти порции энергии не одинаковы по
72
величине. Их величина определяется разницей между редокс - потенциалами двух соседних переносчиков. Если эта разница небольшая, то энергии выделяется мало - она рассеивается в виде тепла. Но на нескольких стадиях ее достаточно, чтобы синтезировать макроэргические связи в молекуле АТФ. Такие участки называются пунктами сопряжения тканевого дыхания и окислительного фосфорилирования, они соответствуют расположению комплексов I, III и IV.
Значит, на каждую пару атомов водорода, отнятых от субстрата (НАДН2) – главная дыхательная цепь, возможен синтез 3-х молекул АТФ; от ФАДН2 (сокращенная дыхательная цепь) – 2-х молекул АТФ.
Для количественной оценки эффективности сопряжения тканевого дыхания и окислительного фосфорилирования вычисляют коэффициент P/O. Он показывает, сколько молекул неорганического фосфата присоединилось к молекулам АДФ в расчете на один атом кислорода. Как правило, если донором протонов выступает НАДН2, то
Р/О = 3, если ФАДН2, то Р/О = 2.
Регуляция окислительного фосфорилирования осуществляется на трех уровнях: 1) на уровне количества субстратов окисления; 2) на уровне количества кислорода в митохондриях; 3) на уровне отношения АТФ/АДФ + Н3РО4
Основным регулирующим фактором является соотношение АТФ/АДФ. Зависимость митохондриального окисления (дыхания) от концентрации АДФ получила название дыхательного контроля. Если клетка находится в состоянии метаболической активности, что сопровождается затратой АТФ, то накапливается АДФ, который, в свою очередь, активирует процессы биологического окисления и синтеза АТФ.
Разобщители тканевого дыхания и окислительного фосфорилирования не прекращают процессов окисления, но снижают синтез АТФ. Дыхательная цепь работает, а АТФ при этом образуется меньше, чем в норме. Это становится возможным по причине того, что такие вещества способны образовывать собственные протонные каналы во внутренней мембране митохондрий, поэтому часть протонов проходит через них, минуя каналы АТФ-синтетазы. Подобным действием обладают гормоны щитовидной железы – тироксин и трийодтиронин. При состояниях, сопровождающихся гиперфункцией щитовидной железы (Базедова болезнь), больным не хватает энергии АТФ, компенсаторно они много едят (нужно большое количество субстра-
73

тов для окисления), но при этом теряют в весе (большая часть энергии выделяется в виде тепла).
Виды фосфорилирования. Понятие о субстратном и окислительном фосфорилировании. АТФ - универсальный источник энергии в клетке и ее использование в процессах жизнедеятельности.
Синтез АТФ носит название фосфорилирования и описывается уравнением:
АДФ + Н3РО4 = АТФ + Н2О.
Эта реакция происходит при условии обеспечения энергией в количестве не менее 32 кДж/моль.
Если источником этой энергии является транспорт электронов по дыхательной цепи внутренней мембраны митохондрий, говорят об
окислительном фосфорилировании. Это главный путь синтеза АТФ в аэробных клетках.
Если источником энергии является гидролиз макроэргической свя-
зи субстрата, говорят о субстратном фосфорилировании. Такой ме-
ханизм имеет место в цитозоле и митохондриях и может происходить
ванаэробных условиях.
Урастений существует фотосинтетическое фосфорилирование в
хлоропластах. Источником энергии в данном случае являются кванты солнечного света.
Вэнергетическом обеспечении клетки важнейшую роль играет АТФ. Это макроэргическое соединение. Как известно, к макроэргическим относятся соединения, при гидролизе которых высвобождается не менее, чем 5 ккал/моль.
Две фосфоангидридные связи в молекуле АТФ являются макроэргическими. Свободная энергия гидролиза каждой из них равна 7,3 ккал/моль (32 кДж/моль). Это так называемая клеточная энергетическая валюта.
74
АТФ используется клетками для процессов биосинтеза (анаболические реакции), активации многих молекул (глюкоза, глицерин), выполнения механической работы, переноса веществ через мембраны, обеспечивает точную передачу генетической информации и др. При этом АТФ может гидролизоваться двумя способами:
1)АТФ + Н2О = АДФ + Н3РО4 + энергия;
2)АТФ + Н2О = АМФ + Н4Р2О7 + энергия.
Основные пути потребления кислорода в реакциях биологического окисления - оксидазный, пероксидазный, оксигеназный и пероксидное окисление ненасыщенных жирных кислот. Токсичность кислорода, регуляторы свободно-радикального окисления в клетках. Антиоксиданты как лекарственные препараты.
Организм человека функционирует в аэробных условиях: 90% энергии он получает при участии кислорода. Все реакции, идущие с потреблением О2, можно разделить на 4 типа:
1. Оксидазный путь S•Н2 + 1/2О2→S+Н2О
Прoдуктами реакции являются oкисленный субстрат и вода. Эти реакции лoкализованы во внутренней мембране митохондрий (дыхательная цепь), и кислoрод расходуется на образoвание энергии.
2. Пероксидазный S•Н2 + О2→S+Н2О2
Прoдуктами реакции являются oкисленный субстрат и перoксид водорода. Ферменты, катализирующие такие реакции нахoдятся в специальных органoидах клетки – пероксисoмах. Кислорoд в реакциях второго типа испoльзуется на oкисление ряда прирoдных водoрастворимых соединений: аминокислот, биогенных аминов и др. Образующийся пероксид водорода тoксичен для oрганизма, однако, в лейкoцитах он испoльзуется для обезвреживания болезнетворных бактерий.
3.Оксигеназный. Эти реакции катализируются монooксигеназами, кoгда один атoм кислорoда включается в oкисляемый субстрат или диоксигеназами – встраивается два атома кислoрода в процессе микросoмального окисления. Данный путь потребления кислорода испoльзуется при окислении природных липофильных органических соединений, а также при обезвреживании лекарств и ядов.
4. Пероксидное окисление ненасыщенных жирных кислот про-
текает по схеме: RH + О2 → RООН. Прoдуктами oкисления являются гидрoпероксиды липидов, спирты, альдегиды и др. Эти реакции прoтекают в мембранах клеток, где имеются ненасыщенные липиды с целью регуляции обнoвления липидов биoлoгических мембран. Ак-
75
тиватoрами пероксидного окисления липидов являются активные формы кислорода: супероксид-ион, гидроксильный радикал, перок- сид-ион, синглентный кислород, котoрые мoгут образoвываться при перенoсе электронов от oкисляемых субстратов на кислород. Данные частицы представляют собой oпасно для жизни клеток вследствие пoвреждений, которые они спoсобны причинить всем классам биомoлекул, особенно белкам и липидам, вызывая перекиснoе oкисление.
В живoй клетке происходит детoксикация перoксида водорода и супероксид-иона при участии природных антиоксидантов (аскорбиновой кислоты, витамина Е, глутатиона) и специальных ферментов (супероксиддисмутазы, каталазы, глутатионпероксидазы).
Общие представления о фотосинтезе.
Фотосинтез – это образование клетками высших растений, водорослей некоторыми бактериями органического вещества при участии энергии света. Фотосинтез происходит в специализированных органеллах растительных клеток – хлоропластах, содержащих поглощающие свет пигменты.
В основе фотосинтеза лежит окислительно-восстановительный процесс, в котором электроны переносятся от донора-восстановителя (Н2О) к акцептору (СО2) с образованием восстановленных соединений (углеводы) и выделением О2. Фотосинтез протекает в две стадии: первая – световые реакции, идущие только тогда, когда растение освещено, а вторая – темновые реакции, которые могут происходить как в темноте, так и на свету. Четыре основных события световой стадии: фотосинтетическое возбуждение хлорофилла, фотоокисление воды (фотолиз), восстановление НАДФ+ до НАДФН2, синтез АТФ. Реакции темновой стадии сводятся к ферментативной утилизации СО2 и превращению его в углеводы, ходе этого процесса используются АТФ и НАДФ2, которые образовались в световую фазу.
I.Вопросы для самоподготовки:
1.Расскажите о структурной организации митохондрий.
2.В чем заключается основная функция митохондрий?
3.Назовите основные фазы извлечения энергии из питательных веществ.
4.Дайте характеристику работы пируватдегидрогеназного комплекса
(ПДК).
5.Напишите суммарную реакцию ПДК.
6.Реакции цикла Кребса, их характеристика.
76
7.Назовите биологические функции цикла Кребса.
8.Что такое «тканевое дыхание», «дыхательная цепь»?
9.Представьте схему работы дыхательной цепи в митохондриях и назовите ее основные комплексы.
10.Расскажите о механизме образования электрохимического потенциала в дыхательной цепи митохондрий.
11.Что такое «окислительное фосфорилирование»?
12.Пункты сопряжения дыхания и фосфорилирования.
13.Регуляция окислительного фосфорилирования.
14.Перечислите основные пути потребления кислорода.
15.Что такое активные формы кислорода и в каких реакциях они образуются?
16.Назовите способы антиоксидантной защиты организма.
17.Что такое фотосинтез?
18.Какие основные процессы протекают в световую стадию?
II. Тестовые вопросы для самоконтроля:
1. Пируватдегидрогеназный комплекс локализован в ...
а) лизосомах б) митохондриях
в) эндоплазматическом ретикулуме г) ядре клетки
2. При окислении пирувата происходит образование… а) НАДН2; ацетил - КоА; СО2 б) ФАДН2; сукцинил - КоА; СО2
в) Н2О; АТФ г) НАДФН2
3. В цикле Кребса образуются …
а) 3 НАДН2; ФАДН2 ; АТФ б) ацетил - КоА; НАДН2; СО2
в) Н2О; АТФ; глюкоза г) НАДФН2 , СО2
4. Водороддонорная функция цикла Кребса заключается в… а) наработке водорода для дыхательной цепи б) наработке НАДФН2 в) образовании воды
г) образовании ацетил – КоА 5. Цитохромы способны переносить …
а) только протоны б) протоны и электроны
77
в) только электроны г) атомы кислорода
6. Транспорт протонов ферментами дыхательной цепи происходит в направлении …
а) из межмембранного пространства митохондрий в цитозоль б) из цитозоля в межмембранное пространство митохондрий
в) из цитоплазмы в полость гладкого эндоплазматического ретикулума г) из матрикса митохондрий в межмембранное пространство
7. Протонный потенциал (ΔµН+) состоит из двух компонентов - … а) водороддонорного и химического б) осмотического и электрического в) физического и концентрационного
г) механохимического и конформационного 8. Образование АТФ из АДФ и Фн с использованием энергии, заключенной в субстрате, называется …фосфорилированием
а) окислительным б) субстратным
в) фотосинтетическим г) химическим
9. Если донором протонов для дыхательной цепи является НАДН2, то возможно образование ... молекул АТФ
а) 1 б) 2 в) 3 г) 4
10. Обратное движение протонов в матрикс возможно при участии
специального фермента...
а) Са2+- АТФазы
б) АТФ-синтетазы в) Nа+,К+-АТФазы г) Nа+,К+-обменника
11. Какой компонент не входит в состав комплекса III ? а) цитохром в б) цитохром с
в) FeS
г) НАДН2
78
12. Какой путь потребления кислорода используется при окислении природных липофильных органических соединений, а также при обезвреживании лекарств и ядов?
а) оксидазный б) оксигеназный
в) пероксидазный г) глюкозоксидантный
13. К антиоксидантам относится … а) метионин б) глюкуроновая кислота в) витамин Е
г) НАДФН2 14. Фермент, участвующий в освобождении клетки от избытка перок-
сидов, называется … а) альдегиддегидрогеназа
б) моноаминооксидаза в) глутатионпероксидаза г) нитратредуктаза
15. Какие из веществ не образуются в процессе фотосинтеза? а) АТФ б) углеводы
в) О2 г) липиды
III. Контрольные задания для самостоятельной работы:
А. Решите следующие ситуационные задачи:
1. Обсудите ситуацию: «У больного в крови увеличено содержание ПВК». Какие метаболические процессы могут привести к накоплению ПВК в организме?
2.В эксперименте на животных исследованием активности ферментов цикла Кребса установлено, что в митохондриях печени под действием некоторых токсических веществ падает уровень оксалоацетата. Какие реакции цикла Кребса при этом нарушаются?
3.Организуйте деловую игру на тему: «Разобщители тканевого дыхания и окислительного фосфорилирования». Для этого вначале изучите хемиосмотическую теорию сопряжения Митчела. Затем назовите примеры физиологических процессов, где тканевое дыхание «отключено» от окислительного фосфорилирования. Подготовьте краткие сообщения о возможных видах разобщителей в организме.
79
Б. 1. Показать в виде схемы пути использования АТФ в организме.
2. Выписать формулы общих ключевых метаболитов и показать в виде схемы возможные пути их образования.
Частный модуль 1.5. Обмен углеводов
После изучения частного модуля 1.5. студент должен:
Знать: общую схему источников и путей расходования глюкозы в организме. Аэробный и анаэробный пути распада глюкозы: последовательность реакций, биологическую роль и энергетическую ценность. Распад и биосинтез гликогена, регуляция и биологическая роль. Глюконеогенез: источники, обходные пути, биологическую роль. Пентозофосфатный путь превращения глюкозы, его роль. Наследственные нарушения обмена моносахаридов и дисахаридов: галактоземию, непереносимость фруктозы, непереносимость дисахаридов. Гликогенозы и агликогенозы.
Уметь: определять содержание глюкозы в крови; оценивать информативность биохимических показателей анализа крови и мочи при патологических состояниях (сахарный диабет); самостоятельно находить достаточное количество информации для решения ситуационных задач.
Владеть: навыками расчета энергетической ценности глюкозы, фруктозы, галактозы в аэробных и анаэробных условиях; способностью самостоятельно проводить химический эксперимент и интерпретировать его результаты.
Общая характеристика и классификация углеводов. Полисахариды. Свойства и отдельные представители полисахаридов, биологические функции.
Углеводами называют органические соединения, большая часть которых (но не все!) имеют общую формулу Сn(Н2О)m.
Классификация углеводов.
Моносахариды
Моносахаридами называют углеводы, которые не могут быть гидролизованы до более простых форм. К триозам (1) относят глицеральдегид, дигидроксиацетон; к тетрозам (2) – эритроза; к пентозам
(3) – рибоза, ксилоза, рибулеза; к гексозам (4) – глюкоза, галактоза, фруктоза и др.
80