

Nikolls_-_Ot_neyrona_k_mozgu
.pdfГлава 6. |
Ионные |
|
механизмы |
потенциала |
действия |
|
123 |
tance in the giant axon of Loligo. J. Physiol I16: 497-506.
о Hodgkin, A. L., and Huxley, A.F. 1952. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 117: 500-544.
Цитированная литература
1. |
Overton, Ε. 1902. |
Pflugers |
|
Arch. 92: 346-386. |
|
|
|
|||
2. |
Hodgkin, A. L., |
and Huxley, A. |
F. 1939. Nature |
|
144:710 |
-711. |
4» |
|||
3.Hodgkin, A. L., and Katz, B. 1949. /. Physiol. 108: 37-77.
4.Keynes, R.D., and Lewis, P.R. 1951. / Physioi. 114: 151-182.
5.Hodgkin, A. L., Huxley, A. F., and Katz, B. 1952. J. Physiol. 116:424-448.
6.Hodgkin. A. L., and Huxley, A. F. 1952. /. Physiol. 116:449-472.
7.Hodgkin, A. L., and Huxley, A. F. 1952. /. Physioi. 116: 473-496.
8.Hodgkin, A. L., and Huxley, A. F. 1952. /. Physiol 116:497-506.
9.Hodgkin, A. L., and Huxley, A. F. 1952. /. Physioi. 117: 500-544.
10.Marmont, G. 1940. J.Cett. Сотр. Physiol. 34: 351-382.
11.Cole, K. S. 1968. Membranes, Ions and Impulses. University of California Press, Berkeley.
12.Kao, C.T. 1966. Pharmacol. Rev. 18: 977-1049.
13.Narahashi, T., Moore, J. W., and Scott, W. R. 1964. /. Gen. Physiol. 47: 965-974.
14.Hille, B. 1970. Prog. Biophys. Mol. Blol. 21: 1-32.
Biophys. Mol. Blol. 21: 1-32.
15.Armstrong, СМ., and Hille, В. 1972. / Gen. Physiol. 59: 388-400.
16.Armstrong, C. M., and Bezanilla, F. 1974. /. Gen. Physiol. 63: 533-552.
17.Keynes, R. D., and Rojas, E. 1974. /. Physiol. 239: 393-434.
18.Armstrong, С. М. 1981. Physiol. Rev. 61: 644-683.
19.Sigworth, F.J., and Neher, Ε. Ι980. Nalure 287: 447-449.
20.Sigworth, F.J. 1994. Q.Rev. Biophys. 27: 1-40.
21.Stiihmer, W.. et al. 1989. Nature 239: 597-603.
22.Papazian, D. M., et al. 1991. Nature 349: 305-349.
23.Yang, N.. George, A. L., and Horn, R. 1996. Neuron 16: 113-122.
24.Larsson, H.P., et al. 1996. Neuron 16: 387-397.
25.Armstrong, C. M., Bezanilla, F., and Rojas, E. 1973. /. Gen. Physiol. 62: 375-391.
26.Armstrong, C. M., and Bezanilla, F. 1977. /. Gen. Physiol. 70: 567-590.
27.West, J.W., et al. 1992. Proc. Nail. Acad. Sci USA 89: 10910-10914.
28.Kallenberger, S., et al. 1997. /. Gen. Physiol. 109: 589-605.
29.Kalienberger, S., et al. 1997. /. Gen. Physiol. 109: 607-617.
30.Catterall, W.A. 1980. Annu. Rev. Pharmacol. Toxi-col. 20: 15-43.
31.Hoshi, T., Zagotta, W. K, and Aldrich,
W. K, and Aldrich, R.W. 1990. Science 250: 533-550.
R.W. 1990. Science 250: 533-550.
32.Zagotta, W. N., Hoshi, T., and Aldrich, R. W. 1990. Science 250: 568-571.
33.Hoshi, T.W.,
T.W., Zagotta, N.. and Aldrich, R.W. 1991. Neuron 7: 547-556.
Zagotta, N.. and Aldrich, R.W. 1991. Neuron 7: 547-556.
34.Choi, K. L, Aldrich, R.W, and Yellen, G. I99I. Proc. Natl. Acad. Sci. USA 88: 5092-5095.
35.Keynes, R. D. 1990. Proc. R. Soc. Land. В 240: 425-432.
36.Keynes, R. D., and Elinder, F. 1998. Proc. R. Sac. Lond. В 265: 263-270.
37.Bean, B.P. 1981. Biophys. J. 35: 595-614.
38.Aldrich, R. W, and Stevens, C. F. 1983. Cold Spring Harb. Symp. Quant. Biol. 48: 147-153.
39.Hille, B. 1992. Ionic Channels of Excitable Membranes, 2nd Ed. Sinauer Associates, Sunderiand, MA, Chapter 18.
40.Patlach, J. 1991. Physiol. Rev. 71: 1047-1080.
41.Aldrich, R. W, and Stevens, C. F. 1987. /. Neurosci. 7:418-431.
42.Cota, G., and Armstrong, C. M. 1989. /. Gen. Physiol. 94: 213-232.
43.Conti, F., and Stuhmer, W. 1989. Eur. Biophys. J. 17: 53-59.
44.Safronov, V. В., and Vogel, W. 1995. /. Physiol. 487: 91-106.
45.Levinson, S.R., and Meves, H. 1975. Phil. Trans. R. Soc. Lond. В 270: 349-352.
46.Waxman, S. G., et al. 1989. Proc. Natl. Acad. Sci. USA 86: 1406-1410.
47.Ritchie, J. M. 1986. Ann. N. Y. Acad. Sci. 479: 385-401.
48.Beam, K. G., Caldwell, J. H., and Campbell, D.T. 1985. Nature 313: 588-590.
49.Aimers, W, Stanfield, P., and Stuhmer, W. 1983. /. Physiol. 336: 261-284.
50.Lanno, I., Webb, C. K., and Bezanilla. F. 1988. /. Gen. Physiol. 92: 179-196.
51.Chiu, S. Y, et al. 1979. /. Physiol. 292: 149-166.
52.Mathie, A., Wooltorton, J. R., and Watkins, C. S. 1998. Gen. Pharmacol. 30: 13-44.

124 |
Раздел II. Передача |
|
информации |
|
в нервной |
|
системе |
53.Vergara, С., et al. 1998. Cuir. Opin. Neurobiol. 8: 321-329.
54.Meech, R.W. 1974. J.Physial. 237: 259-277.
55.Partridge, L. D., and Thomas, R. C. 1976. /. Phy-siol. 254: 551-563.
56.Martin, A. R., and Dryer, S. E. 1989. Q. J. Exp. Physiol. 74: 1033-1041.
57.Koh, D-S., Jonas, P., and Vogel, W. 1994. /. Phy-siol. 479 (Pt.
(Pt. 2): 183-197.
2): 183-197.
58.Fait, P., and Ginsberg, B.L. 1958. J. Physiol. 142: 516-543.
Physiol. 142: 516-543.
59.Hagiwara, S., and Byerly, L. 1981. Anna. Rev. Neurosci. 4: 69-125.
60.Hagiwara, S. 1983. Membrane Potential-Dependent Ion Channels in Ceil Membrane. Phylogenetic and
Developmental Approaches. Raven, New York.
61.Llinas, R., and Sugimori, M. 1980. /. Physiol. 305: 197-213.
62.Ross, W. N., Lasser-Ross, N., and Werman, R. 1990. Proc. R. Soc. Land. В 240: 173-185.
63.Frankenhaeuser, В., and Hodgkin, A. L. 1957. J. Physiol. 137:218-244.
Physiol. 137:218-244.
64.Miller, J. A., Agnew, W. S., and Levinson, S. R. 1983. Biochemistry 22: 462-470.
65.Hille, B. 1968. J.Gen. Physiol. 51: 221-236.
Глава 7 Нейроны как проводники электричества
Потенциал действия продвигается вдоль аксона за счет продольного распространения тока. Каждый участок мембраны, генерируя импульс по принципу «все или ничего», своей деполяризацией возбуждает соседний участок. Для понимания механизма распространения импульса, а также синаптической передачи и интеграции, необходимо уяснить принцип пассивного распространения импульсов вдоль нерва.
По мере своего продвижения вдоль аксона или дендрита ток теряет свою силу. Снижение величины тока зависит от ряда причин, в первую очередь от диаметра и свойств мембраны нервного волокна. Продольное распространение тока тем больше, чем больше диаметр волокна и чем выше сопротивление мембраны. Емкостные свойства мембраны влияют на временной ход электрических сигналов, а также на распространение тока. Чтобы оценить, на какое расстояние способен распространиться подпороговый потенциал, необходимо знать геометрию и свойства мембраны нейрона и, кроме того, временной ход изменения потенциала.
Аксоны многих типов нервных клеток у позвоночных покрыты оболочкой из миелина, обладающего высоким сопротивлением и малой емкостью. Миелиновая оболочка играет роль изоляционной обмотки провода и заставляет ток перемещаться вдоль мембраны. При этом ток быстро перескакивает от одного перехвата Ранвье (короткого участка мембраны, лишенного миелина) до другого, и скорость проведения при этом возрастает. Миелинизированные волокна встречаются в тех областях нейронной сети, где скорость проведения играет важную роль.
Электрические сигналы способны также передаваться с одного нейрона на другой в местах тесного контакта между ними, называемых щелевыми контактами. Ток в таких соединениях протекает по особым каналам, коннексонам.
§ 1. Пассивные электрические свойства нервных и мышечных мембран
Особенности проницаемости нейрональных мембран и то, как они способствуют генерации потенциала действия, обсуждались в предыдущих главах. В данной главе речь пойдет главным образом о том, как токи распространяются вдоль нервного волокна и образуют локальные разности потенциалов.
Пассивные электрические свойства нейронов, а именно, сопротивление и емкость мембраны, а также сопротивление цитоплазмы, играют важнейшую роль в нейрональной сигнализации. В органах чувств эти свойства являются связующим звеном между сенсорным стимулом и генерацией импульса; на уровне аксона они позволяют импульсу распространиться; на уровне синапсов они определяют способность постсинаптического нейрона складывать и вычитать синаптические потенциалы, возникающие на многочисленных синаптических входах, будь то вблизи тела клетки или на самых отдаленных дендритах. Для понимания этих процессов нужно знать принципы распространения электрических сигналов вдоль нейронных отростков. В данной главе речь пойдет главным образом о нервных волокнах с постоянным диаметром на всем своем протяжении, т. е. о цилиндрических проводниках. Кроме того, для данного описания мы примем, что в отсутствие регенерации мембрана действительно пассивна, т. е. изменения потенциала, не достигающие порогового уровня, не активируют потенциалзависимых проводимостей и не изменяют тем самым сопротивления мембраны. Данные принципы применимы и к более сложным структурам, таким как разветвления аксонных окончаний или дендритные ветвления с неоднородными электрическими свойствами 1· 2). Роль подобных структур в функционировании нервной системы весьма велика, однако количественное описание их электрических свойств потребовало бы более сложного анализа.
-
126 |
Раздел II. Передача информации в нервной системе |
Кабельные свойства нервных и мышечных волокон
Цилиндрическое нервное волокно состоит из тех же компонентов, что и подводный электрический кабель: из стержневого проводника и изоляционной оболочки, окруженной проводящей средой. Тем не менее, количественное отличие этих двух систем весьма велико. Стержень кабеля обычно сделан из меди или металла с очень высокой проводимостью, в то время как оболочка сделана из пластика или других материалов с очень высоким сопротивлением. Кроме того, оболочка обычно бывает довольно толстая и потому обладает низкой емкостью (приложение А). Напряжение, приложенное к такому проводу, способно передаться на значительное расстояние благодаря тому, что сопротивление меди мало, как незначительны и потери через оболочку. Содержимое нервного волокна представляет собой раствор солей, по концентрации похожий на внеклеточную среду и, в отличие от меди, обладающий плохой проводимостью. Мембрана клетки, в свою очередь, не является хорошим изолятором и обладает высокой емкостью ввиду своей малой толщины. Напряжение, приложенное к нервному волокну, не распространяется на значительное расстояние по двум причинам: 1) проводимость содержимого волокна мала, следовательно, сопротивление току велико; 2) ток, протекающий вдоль цитоплазмы, рассеивается благодаря утечке сквозь мембрану, не обеспечивающую достаточной изоляции.
Анализ тока в кабеле был начат лордом Кельвином применительно к трансатлантической телефонной связи и усовершенствован Оливером Хевисайдом. В конце XX в. Хевисайд впервые учел значимость утечки тока через изоляционную оболочку, эквивалентную клеточной мембране, а также внес множество важных дополнений в кабельную теорию, в том числе определил понятие импеданса. Кабельная теория была впервые использована для нервных волокон Ходжкиным и Раштоном3), которые экспериментально измерили распространение потенциала действия в аксоне омара с помощью внеклеточных электродов. Позднее для подобных измерений в целом ряде нервных и мышечных волокон использовались внутриклеточные электроды.
В данном разделе рассматривается зависимость продольного распространения тока от сопротивления мембраны и аксоплазмы. Основным правилом здесь является закон Ома: ток i, проходя через сопротивление г, создает напряжение ν = ir (приложение А). Ниже в этой главе мы рассмотрим также влияние мембранной емкости на величину и временной ход продольного распространения тока.
Ток в кабеле
Для лучшего понимания принципов прохождения тока по кабелю представим себе, как распространяется тепло вдоль металлического прута в теплоизолирующей оболочке, который помещен в проводящую среду (например, в воду). Если нагревать прут с одного конца, тепло распространяется вдоль прута и, по мере своего распространения, частично рассеивается в окружающую среду, проникая через оболочку. Чем дальше от нагреваемого конца, тем ниже температура; по мере снижения температуры снижается и скорость рассеяния тепла. При условии, что окружающая среда обладает высокой теплопроводностью, расстояние, на которое распространится теплота, будет зависеть главным образом от двух параметров: 1) от теплопроводности прута и 2) от изолирующих свойств оболочки.
Протекание тока в кабеле происходит похожим образом. Напряжение, приложенное к одному концу кабеля, вызывает протекание тока по стержню в продольном направлении, который частично теряется сквозь оболочку в окружающую среду. Чем дальше от конца, к которому приложено напряжение, тем меньше ток. Расстояние, на которое распространится ток, будет зависеть от проводимости стержня и от того, насколько эффективно оболочка предотвращает потери тока. Оболочка с низким сопротивлением пропустит весь ток в окружающую среду. Оболочка с более высоким сопротивлением позволит току распространиться на большее расстояние.
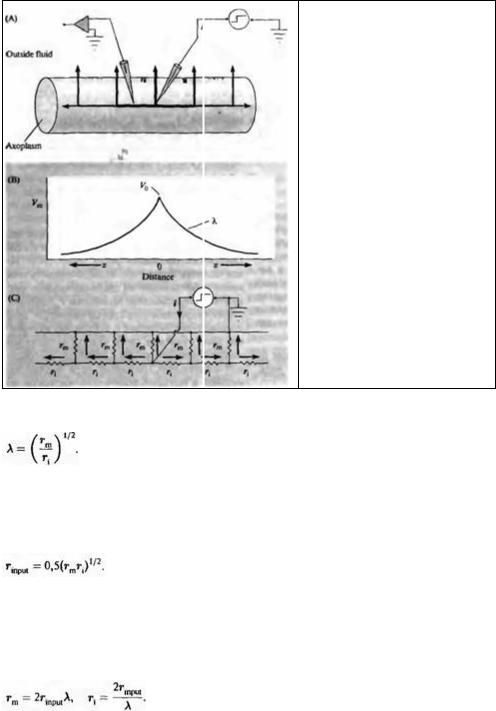
Ток в аксоне переносится ионами: при инъекции тока в нервное волокно (например, в аксон омара) через микроэлектрод, как показано на рис. 7.1 А, инъецированные положительные заряды будут отталкивать другие катионы и притягивать анионы. Самый распространенный из небольших ионов внутри клетки — это калий, который, следовательно, переносит наибольшее количество тока через мембрану. Ток протекает в продольном

Глава 7. Нейроны как проводники электричества |
127 |
направлении вдоль аксона, и по мере продвижения часть его теряется благодаря перемещению ионов через мембрану. В мембране с низким сопротивлением и большой ионной проводимостью большая часть тока потеряется до того, как он успеет переместиться на скольнибудь значительное расстояние. При более высоком сопротивлении мембраны ток распространится вдоль аксона на большее расстояние, прежде чем рассеяться в окружающую среду.
Входное сопротивление и постоянная длины
На рис. 7.1А представлен ток, протекающий через различные участки мембраны вдоль аксона на разном расстоянии от отводящего электрода. Толщина каждой стрелки приблизительно соответствует величине тока в каждом отдельно взятом участке. По закону Ома падение напряжения на данном расстоянии от электрода пропорционально мембранному току на том же расстоянии. Возникает два вопроса:
1)Каково будет изменение напряжения на электроде при данном количестве тока, проходящем через электрод?
2)Как далеко это изменение напряжения распространится вдоль волокна?
Чтобы ответить на эти вопросы, достаточно измерить изменение напряжения с помощью еще одного электрода, вставляемого в волокно на различном расстоянии от первого электрода, как показано на рис. 7.1 А. Результаты подобных измерений показаны на рис. 7.1В. Изменение напряжения имеет наибольшее значение в месте инъекции тока и постепенно снижается по мере удаления от него.
Спад величины изменения напряжения носит экспоненциальный характер, так что потенциал (Vz) при данном значении расстояния x равен:
Максимальный потенциал V0 пропорционален величине инъецируемого тока. Коэффициент
пропорциональности называется входным сопротивлением волокна, rinput. Он равен среднему сопротивлению, встречаемому ионным током, протекающим через аксоплазму и мембрану во внеклеточную среду. Таким
образом, если величина инъецированного тока равна i, то
Постоянная длины волокна λ — это расстояние, на котором потенциал снижается в е раз (до 37% от начального значения). Эти два параметра, rinput и λ, определяют величину изменения потенциала в ответ на инъекцию тока, а также расстояние, на которое это изменение распространится вдоль волокна.
Сопротивление мембраны и продольное сопротивление
Кабель можно представить себе в виде цепочки элементов сопротивления, rm и ri, соединенных последовательно (рис. 7.1C). Такая цепочка получится, если вообразить себе аксон разрезанным на цилиндрические сегменты. Сопротивление мембраны rm соответствует сопротивлению стенки цилиндра; продольное сопротивление ri составляет внутреннее сопротивление вдоль участка аксоплазмы между серединой данного цилиндра и серединой следующего. Поскольку нерв обычно помещают в большое количество жидкости, то внеклеточное продольное сопротивление можно принять за ноль. Это допущение не всегда верно для центральной нервной системы, в которой аксоны и дендриты нейронов, а также глиальные клетки (см. главу 8) упакованы настолько плотно, что протекание внеклеточного тока ограничено. В случае рассматриваемого опыта такое допущение справедливо и служит для максимального упрощения математических выражений. Длина цилиндрических сегментов может быть любой, однако rm и ri ( принято указывать из расчета на участок аксона длиной 1 см. Сопротивление мембраны rm выражается в омах, умноженных на сантиметр (Ом см). Величина может показаться странной, но она объясняется тем, что сопротивление убывает с увеличением длины волокна, а значит, с увеличением количества каналов, через которые теряется ток. Таким образом, сопротивление данного участка мембраны аксона в омах равняется сопротивлению сантиметрового участка (rm в Ом · см), деленному на длину данного участка (в см). Постоянная ri; выражается, как и следовало ожидать, в омах, деленных на сантиметр (Ом/см).
128 |
Раздел II. Передача информации в нервной системе |
Рис. 7.1. Токи в мембране аксона. (А) Ток (i), протекающий в мембране, в ответ на инъекцию постоянного тока через микроэлектрод. (В) Потенциал (Vm) как функция расстояния (x) от места инъекции тока. (С) Эквивалентная электрическая схема, в которой сопротивление внеклеточной среды принято за ноль, а емкость мембраны не учитывается.
Fig. 7.1. Pathways for Current Flow in an axon. (A) Current flow (i) across the membrane produced by a steady injection of current from a microelectrode. Thickness of the arrows indicates current density at various distances from the point of injection. The second electrode (left) records membrane potential at various distances from the current electrode. (B) Potential (Vm) measured along the axon as a function of distance (x) from the point of current injection. Decay of voltage is exponential with a length constant λ. (С) Equivalent electrical circuit assuming zero resistance in the external fluid and ignoring membrane capacitance, ri = longitudinal resistance of axoplasm per unit length; rm = membrane resistance of a unit length.
Расчет сопротивления мембраны и внутреннего сопротивления
Постоянная длины волокна зависит как от rm, так и от ri:
Величина измеряется в сантиметрах и отвечает интуитивному представлению о том, что расстояние, на которое распространяется изменение потенциала, должно возрастать с увеличением сопротивления мембраны (которое препятствует потере тока за счет утечки) и, напротив, должно снижаться с увеличением внутреннего сопротивления (которое затрудняет протекание тока вдоль содержимого волокна).
Точно так же входное сопротивление зависит от обоих параметров:
Выражение измеряется в должных единицах (Ом) и подразумевает возрастание входного сопротивления с увеличением как одного, так и другого параметров. Фактор 0,5 объясняется тем, что аксон простирается в двух направлениях от места инъекции тока. Каждая половинка обладает входным сопротивлением, равным (rmri) 0,5.
Зная эти выражения, можно получить параметры сопротивления мембраны и аксоплазмы из опытов, подобных показанному на рис. 7.1. Измерив экспериментально rinput и λ, можно путем простых преобразований получить выражения для rm и ri:
Удельное сопротивление
Рассчитанные значения rm и ri характеризуют сопротивление цилиндрического сегмента аксона длиной 1 см. Однако, они не предоставляют точной информации о сопротивлении самой мембраны, а также аксоплазмы, поскольку последние зависят от размера волокна. При равных прочих условиях, сантиметровый отрезок тонкого волокна должен обладать более высоким сопротивлением мембраны, чем отрезок более толстого волокна
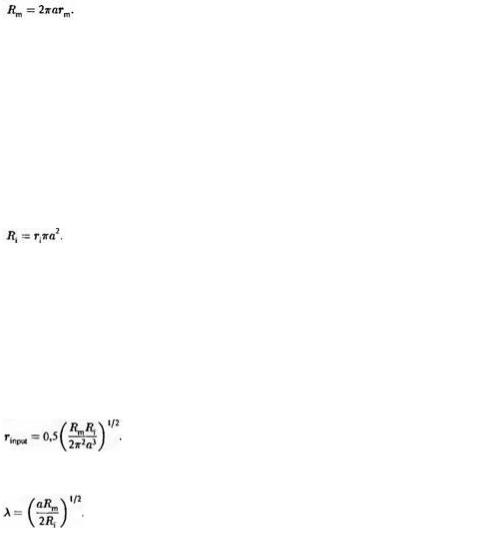
Глава 7. Нейроны как проводники электричества |
129 |
той же длины, поскольку у тонкого волокна площадь поверхности мембраны меньше, чем у толстого. С другой стороны, если в мембране тонкого волокна плотность каналов значительно выше, то сопротивление сантиметрового участка его мембраны может в результате оказаться таким же, как и у толстого волокна. Для того, чтобы сопоставить мембраны между собой, нужно знать величину удельного сопротивления, которое равно сопротивлению мембраны площадью 1 см2 и измеряется в Ом см2. Отрезок аксопа длиной 1 см и радиусом о обладает площадью поверхности 2тго см2. Сопротивление мембраны rm такого отрезка можно получить, разделив Rm на площадь мембраны: rm = Rm/(2πа). Преобразовав выражение, получим
Величина Rm важна тем, что не зависит от геометрических свойств мембраны и потому
позволяет сравнивать между собой мембраны самых разных размеров и форм.
В большинстве типов нейронов Rm определяется главным образом калиевой и хлорной проницаемостями мембраны в покое (глава 5), величина которых зависит от типа клетки. Среднее значение Rm, полученное Ходжкиным и Раштоном на аксоне омара, составляет приблизительно 2000 Ом · см2; данные, полученные в других препаратах, варьировали в диапазоне от 1 000 Ом · см2 в мембранах с большим количеством каналов до 50000 Ом см2 в мембранах со сравнительно небольшим числом каналов.
Удельным сопротивлением (Ri) аксоплазмы называется внутреннее продольное сопротивление участка аксона длиной I см и площадью сечения 1 см2. Эта величина также не зависит от геометрических свойств аксона и определяется тем, насколько свободно ионы способны перемещаться во внутриклеточном пространстве. Чтобы получить Ri из ri, нужно вспомнить, что с увеличением площади сечения цилиндра его продольное сопротивление уменьшается. Следовательно, сопротивление участка аксона длиной 1 см (ri) можно выразить как частное величины Ri и площади сечения: ri = Ri/(πа2). Отсюда получим:
Величина R, измеряется в Ом · см. Для аксона кальмара ее значение составляет приблизительно 30 Ом см при 20° С, что в 107 раз выше, чем удельное сопротивление меди. Величина эта соответствует ожиданиям для ионного состава аксоплазмы кальмара3). У млекопитающих ионная концентрация в цитоплазме ниже, а удельное сопротивление выше, около 125 Ом · см при 37° С. У лягушек, при еще более низкой ионной концентрации, величина удельного сопротивления при 20° С составляет приблизительно 250 Ом · см.
Влияние диаметра кабеля на его характеристики
Как будет влиять диаметр волокна на его кабельные параметры rinput и λ при данных значениях удельного сопротивления аксоплазмы Ri и удельного сопротивления мембраны Rm? Ответ можно получить на основе уравнений, приведенных выше. Начнем с входного сопротивления.
Мы знаем, что rinput = 0,5(rmri)1/2 и rm = Rm/(2πа), ri = Ri/(2πа2).
Преобразовав эти выражения, получим:
Следовательно, с увеличением диаметра волокна о входное сопротивление снижается как 3/2 степень радиуса.
Подобным же образом найдем постоянную длины:
При равных прочих условиях λ возрастает с увеличением радиуса волокна как квадратный корень радиуса. Этим параметром можно пользоваться при сравнении различных волокон между собой, если принять, что удельное сопротивление мембраны равно 2000 Ом см2 в каждом случае. Аксон кальмара длиной I см с удельным внутренним сопротивлением 30 Ом · см должен обладать постоянной длины почти 13 мм. Мышечное волокно лягушки, благодаря своему меньшему диаметру и более высокому удельному внутреннему сопротивлению, при диаметре 50 мкм имеет постоянную длины, равную всего 1,4 мм, а нервное волокно млекопитающих диаметром 1 мкм — всего 0,3 мм.
Обобщим вышесказанное. Кабельные параметры rinput и λ определяют величину сигнала, генерируемого нервным волокном, а также расстояние, на которое этот сигнал способен распространиться. Например.

130 |
Раздел П. Передача информации в нервной системе |
при равных прочих характеристиках, величина возбуждающего синаптического потенциала
(глава 9) будет больше в тонком дендрите (с более высоким rinput), чем в толстом. С другой стороны, в толстом дендрите потенциал будет распространяться на большие расстояния по
направлению к телу клетки, чем в тонком (благодаря большему значению λ). Тем не менее, rinput и λ зависят не только от размера волокна, но и от резистивных свойств цитоплазмы и мембраны. Можно с высокой степенью аккуратности считать, что значение удельного сопротивления цитоплазмы одинаково для любых клеток независимо от класса животного. Удельное сопротивление мембраны, напротив, различно для разных типов клеток и может отличаться в десятки раз.
Емкость мембраны
Мембрана клетки не только проводит ионные токи, но и накапливает заряд на своей внешней или внутренней поверхности (глава 5). С точки зрения теории электричества, разделение зарядов на мембране означает, что мембрана обладает свойствами конденсатора. В обшем виде конденсатор состоит из двух проводящих пластин, отделенных друг от друга изолирующим материалом; в промышленных конденсаторах проводящие пластины обычно сделаны из металлической фольги, а изолирующая прослойка между ними — из пластика. В случае нервной клетки проводниками являются два слоя жидкости, находящиеся по обе стороны мембраны, а сама мембрана играет роль изолирующей прослойки. При зарядке конденсатора от батареи на одной из пластинок накапливается положительный заряд, в то время как на второй пластинке создается равный по величине запас отрицательного заряда. Емкость конденсатора (С) определяется количеством заряда (Q), запасаемым на один вольт потенциала (V), приложенного к пластинам конденсатора: С = Q/V. С измеряется в кулонах, деленных на вольт, т. е. в фарадах (Ф). Чем ближе друг к другу расположены пластины, тем более эффективно конденсатор способен разделять и накапливать заряд. Поскольку толщина мембраны клетки всего 5 нм, она способна накапливать достаточно большой заряд. Обычно емкость мембраны нервных клеток составляет 1 мкФ/см2. Преобразовав выражение, получаем Q = CV. При потенциале покоя —80 мВ, количество избыточного отрицательного заряда на внутренней стороне мембраны составит (1 · 10–6) х (80 ·10–3) = 8· 10–8 кулонов, деленных на см2, что соответствует 5 · 1011 одновалентных ионов (0,8 пмоль) на квадратный сантиметр мембраны.
Величину тока, протекающего внутрь конденсатора или из него, можно подсчитать на основе соотношения заряда и напряжения, учитывая, что ток (i, в амперах) есть скорость изменения заряда во времени, т.е. 1 ампер = (1 кулон)/(1 с). Поскольку Q = C/V, получим:
Скорость изменения заряда на конденсаторе прямо пропорциональна величине тока. Если ток постоянен, то потенциал будет меняться с постоянной скоростью dV/dt = i/C.
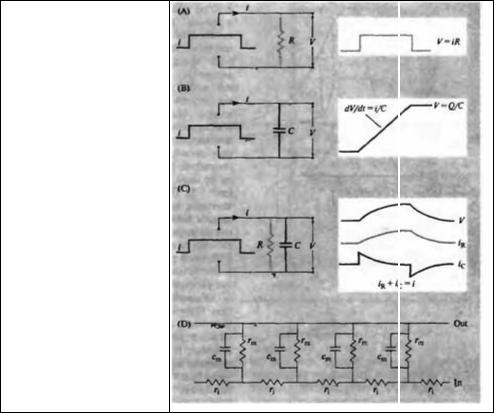
Соотношение тока и напряжения в цепи, содержащей резисторы (сопротивления) и конденсаторы (емкости), соединенные параллельно, проиллюстрировано на рис. 7.2. Прямоугольный скачок тока величиной i, приложенный к резистору (R), создает скачок напряжения на резисторе величиной V = iR (рис. 7.2А). Если тот же скачок тока приложить к конденсатору (С), то напряжение на конденсаторе будет накапливаться со скоростью dV/dt = i/C (рис. 7.2В). Когда эти два элемента, резистор и конденсатор, соединены параллельно (рис. 7.2С), то весь ток пойдет сначала на зарядку конденсатора со скоростью i/С; однако, как только на конденсаторе накопится какой-то заряд, ток потечет и через резистор. По мере нарастания тока, все большая его часть будет проходить через сопротивление, потому что скорость зарядки конденсатора будет постепенно снижаться. В конце концов весь ток будет течь через резистор, создавая на нем потенциал V = iR, a конденсатор будет полностью заряжен. По завершении скачка тока заряд из конденсатора постепенно рассеется на резисторе, а напряжение вернется к нулю.
Постоянная времени
Нарастание и спад потенциала происходит по экспоненциальной кривой, как показано на рис. 7.2В. Фаза роста описывается уравнением:
где t — время от начала импульса. Постоянная времени τ равна произведению RC. Это время, за которое потенциал возрастает
Глава 7. Нейроны как проводники электричества |
131 |
Рис. 7.2. Влияние емкости на временной ход изменения потенциала. (А) Потенциал, создаваемый прямоугольным скачком тока в цепи, содержащей исключительно резисторы. (В) В цепи, состоящей из одних емкостей, скорость изменения потенциала пропорциональна величине прилагаемого тока. (С) Ток в цепи, содержащей как резисторы, так и емкости. (D) Электрическая схема из рис. 7.1С с добавлением удельной емкости мембраны.
Fig. 7.2. Effect of Capacitance on Time
Course of potentials. (A) Potential (V) produced by a rectangular pulse of current (i) in a purely resistive network. Voltage is proportional to, and has the same time course as, the applied current. (B) In a purely capac itative network the rate of change of voltage is proportional to the applied current. (C) In a combined RC network the initial surge of current is into the capacitor (ic); by the end of the pulse all of the current flows through the resistor (iR). Voltage rises to the final value iR exponentially with time constant τ =
RC. After termination of the current pulse, the capacitance discharges through the resistance with the same time constant and ic and iR are equal and opposite. (D) Electrical model of a cable as in Figure 7.1C, but with membrane capacitance per unit length (cm) added.
до 63 % ( 1 - 1/e) своего максимального значения. Спад напряжения тоже экспоненциален, с той же постоянной времени. Ток через резистор, iR, должен изменяться во времени по тому же закону, что и напряжение. Следовательно, на фазе роста ток начинает расти от нуля до своего максимального значения i. Емкостной ток, наоборот, начинается с величины i и спадает до нуля по экспоненте с той же постоянной времени. По завершении импульса, поскольку внешнего источника тока нет, единственным током на резисторе будет ток, генерируемый напряжением на конденсаторе. Следовательно, ток на резисторе равен по величине емкостному току и противоположен ему по направлению, как показано на рисунке.
Вышеописанную цепь, состоящую из параллельно соединенных резистора и конденсатора, можно использовать для описания сферической нервной клетки, аксон и дендриты которой настолько малы, что их вкладом в электрические свойства клеток можно пренебречь. В эквивалентной цепочке для аксона или мышечного волокна как емкость мембраны, так и ее сопротивление распределены по всей длине волокна, как показано на рис. 7.2D. Емкость мембраны на единицу длины ст (измеряемая в мкФ/см) зависит от удельной емкости на единицу площади Сm (в мкФ/см2) по формуле сm = 2πаСт, где α — радиус волокна.
Постоянная времени мембраны сферической клетки или волокна (τm = RтСт) не зависит от размера клетки или волокна. Причина этого в том, что увеличение радиуса (а следовательно, и площади поверхности мембраны) влечет за собой не только увеличение емкости, но и соответствующее снижение сопротивления, так что произведение двух величин не меняется. Поскольку показано, что величина Ст приблизительно одинако-
132 |
Раздел II. Передача информации в нервной системе |
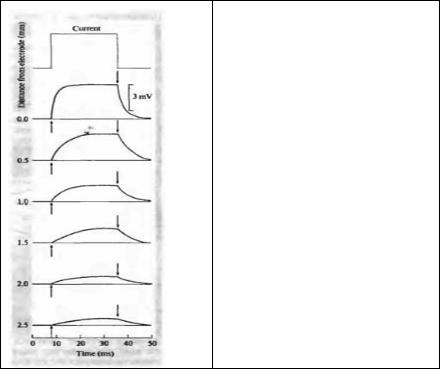
Рис. 7.3. Распространение потенциала вдоль аксона омара, регистрируемое с помощью поверхностного электрода.
Fig. 7.3. Spread of Potential along a lobster axon, recorded with a surface electrode. A rectangular current pulse is applied at 0 mm, producing an electrotonic potential. With increasing distance from the site of current injection, the rise time of the potential change is slowed and the height of the plateau attenuated. (After Hodgkin and Rushton, 1946.)
ва для всех нервных и мышечных волокон (1 мкФ/см2), то величина τ является удобным параметром, характеризующим удельное сопротивление мембраны для данной клетки. Постоянная времени — это третий параметр, который, наряду с входным сопротивлением и постоянной длины, характеризует поведение аксона. Диапазон значений постоянной времени в различных типах нервных и мышечных клеток составляет от 1 до 20 мс.
Емкость в кабеле
Каково влияние постоянной времени на ток в кабеле? Как и в случае простой RC-цепочки (рис. 7.2С), нарастание и спад потенциала в ответ на скачкообразное изменение тока замедляется благодаря наличию конденсатора. Ситуация осложняется тем, что ток течет уже не через один конденсатор, но каждый сегмент цепочки является одним из резистивно-емкостных элементов, взаимодействующих между собой. Благодаря этим взаимодействиям временной ход в отдельном сегменте нельзя описать простой экспонентой, и фазы роста и спада замедляются по мере удаления от точки инъекции тока (рис. 7.3). Поскольку скорость нарастания потенциала зависит от расстояния между отводящим электродом и местом инъекции тока, постоянную времени спада уже нельзя рассчитать на основе простого измерения времени 63 %-ного роста потенциала, за исключением единственной точки вдоль волокна, в которой это расстояние равно постоянной длины.
Обратимся к рассмотрению движения ионов. Если в аксон инъецирован положительный ток, внутриклеточные ионы (главным образом, калия) будут распространяться вдоль волокна. Некоторая часть ионов уйдет на изменение заряда на емкости мембраны, другая часть будет протекать по мембранному сопротивлению. Одновременно с этим процессом будет происходить перемещение отрицательных ионов в противоположном направлении. Постепенно потенциал на мембране достигнет нового устойчивого значения, емкости, распределенные по мембране, будут полностью заряжены до нового уровня потенциала, а через мембрану будет протекать постоянный ионный ток. Время, необходимое для достижения нового устойчивого состояния, определяется постоянной времени.
Еще одно следствие наличия мембранной емкости заключается в том, что короткие сигналы распространяются на более короткие расстояния, чем длительные сигналы. В случае достаточной длительности сигнала, в течение которого потенциал успевает достигнуть своего максимального значения, емкость заряжается полностью, и пространственное распределение потенциала определяется сопротивлениями мембраны и цитоплазмы: Vx = V0ε–x/λ. Для коротких импульсов, таких как синаптический потенциал, ток прекращается еще до того, как емкость успевает полностью зарядиться. Это выражается