

1. Поток вещества
![]() - масса вещества, переносимого через
площадь
- масса вещества, переносимого через
площадь
![]() ,
перпендикулярную движению частиц, в
единицу времени.
,
перпендикулярную движению частиц, в
единицу времени.
2. Плотность потока вещества - масса вещества, переносимого через единицу площади, перпендикулярной движению частиц, за единицу времени.
Молекулы жидкости перескакивают из
одного равновесного состояния в другое.
В одном из равновесных состояний поместим
площадку
![]() и определим, какое число молекул пройдет
через неё из двух ближайших равновесных
состояний
и определим, какое число молекул пройдет
через неё из двух ближайших равновесных
состояний![]() и
и![]() .
.
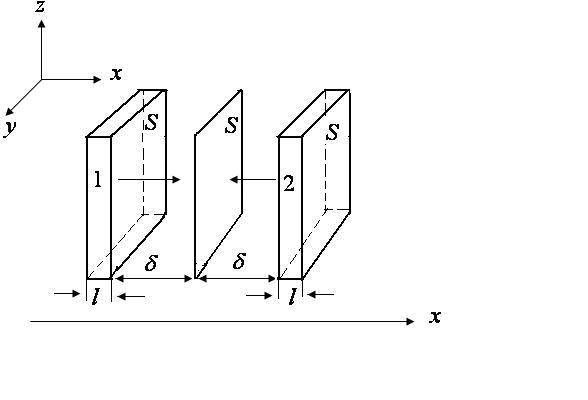
Отложим слева и справа от
![]() расстояния
расстояния![]() (длина свободного пробега молекулы) и
построим параллепипеды
(длина свободного пробега молекулы) и
построим параллепипеды![]() и
и![]() ,
площадью
,
площадью![]() и
толщиной
и
толщиной![]() .
.
Объёмы параллепипедов
![]() .
.
Для упрощения будем считать, что
молекулы движутся с одинаковыми средними
скоростями![]() ,
где
,
где![]() -
время свободного пробега (перескока).
-
время свободного пробега (перескока).
Пусть число молекул в единице объёма
(концентрация) в первом объёме
![]() ,
во втором
,
во втором![]() .
Тогда общее число молекул в первом
объёме
.
Тогда общее число молекул в первом
объёме![]() ,
во втором -
,
во втором -![]() .
.
Молекулы движутся хаотично по всем
трем направлениям:
![]() .
Из них
.
Из них![]() движется в направлении оси
движется в направлении оси![]() ,
,![]() -
-![]() ,
,![]() -
-![]() .
Причем в направлении
.
Причем в направлении![]() ,
например, половина из этой
,
например, половина из этой![]() движется влево, половина - вправо. Поэтому
вдоль оси
движется влево, половина - вправо. Поэтому
вдоль оси![]() от первого параллепипеда к площадке
от первого параллепипеда к площадке![]() движется
движется![]() молекул, а от второго -
молекул, а от второго -![]() .
Т.о. за время
.
Т.о. за время![]() через площадку
через площадку![]() проходит число молекул
проходит число молекул
![]() .
.
Пусть масса одной молекулы
![]() ,
тогда за время
,
тогда за время![]() через
через
площадку
![]() переносится масса вещества
переносится масса вещества
![]() .
.
Время пролета молекулами площадки
![]() параллепипедов
параллепипедов![]() и
и![]() (через центральную оно будет таким же)
(через центральную оно будет таким же)
![]() .
.
Поток вещества через площадку определится как
![]() .
.
Можно показать, что
![]() ,
,
где
![]() градиент концентраций. Поток вещества
тогда запишем как
градиент концентраций. Поток вещества
тогда запишем как
![]()
Через единицу площади переносится поток вещества (плотность потока)
![]() .
.
Введем массовую концентрацию
![]() .
Тогда
.
Тогда
![]() - градиент массовой концентрации.
- градиент массовой концентрации.
Обозначим
![]() и назовем её коэффициентом диффузии, а
также учтем, что суммарная плотность
потока направлена в сторону уменьшения
концентрации, и укажем это напрвавлние
знаком “-“. Получим уравнение Фика:
и назовем её коэффициентом диффузии, а
также учтем, что суммарная плотность
потока направлена в сторону уменьшения
концентрации, и укажем это напрвавлние
знаком “-“. Получим уравнение Фика:
![]() .
.
![]() .
Физический смысл коэффициента диффузии:
он численно равен массе вещества,
переносимого через единичную площадку
за 1 секунду при градиенте концентраций
равном 1.
.
Физический смысл коэффициента диффузии:
он численно равен массе вещества,
переносимого через единичную площадку
за 1 секунду при градиенте концентраций
равном 1.
![]() ,
если
,
если
![]() ,
,
![]() ,
если
,
если
![]()
Лекция 7.
Транспорт веществ через мембрану.
Являясь открытой термодинамической сиситемой, клетка постоянно осуществляет обмен веществом с окружающей средой. Такой обмен возможен благодаря способности клеток пропускать различные вещества через свою оболочку. Эта способность клеток называется проницаемостью.
Перемещение веществ в клетку или из неё в окружающую среду может осуществляться многими способами. В зависимости от того, что является источником энергии для переноса вещестыва, что является силой перемещения, все виды переноса веществ можно разделить на пассивный и активный транспорт.
Пассивный транспорт веществ.
Пассивный транспорт всегда осуществляется за счет энергии, сконцентрированной в каком-либо градиенте. Энергия метаболических процессов клеток (энергия гидролиза АТФ) на этот процесс непосредственно не переносится. Пассивный транспорт всегда идет от более высокого энергетического уровня к более низкому.
Основными градиентами, присущими живым организмам, являются градиенты концентрационные, электрические, осмотические, градиенты гидростатического давления.
В соответствии с этими градиентами имеются следующие виды пассивного транспорта веществ в клетках и тканях: диффузия, осмос, электроосмос, аномальный осмос, фильтрация.
Основным механизмом пассивного транспорта является диффузия- самопроизвольный процесс проникновения вещества из области большей концентрации в область ментшей концентрации в результате теплового хаотического движения.
Перенос незаряженных частиц (атомов и молекул) через мембрану
Очевидно, что при переносе незаряженных частиц единственным градиентом является градиент концентраций. Следовательно, механизм переноса молекул и атомов через мембрану – диффузия, а сам транспорт описывется уравнением Фика:
![]()
Концентрационный градиент клеточной мембраны определить трудно, поэтому лучше использовать более простое уравнение, предложенное Коллендором и Берлундом и которое легко вывести из уравнения Фика.
![]() -
концентрация вещес
-
концентрация вещес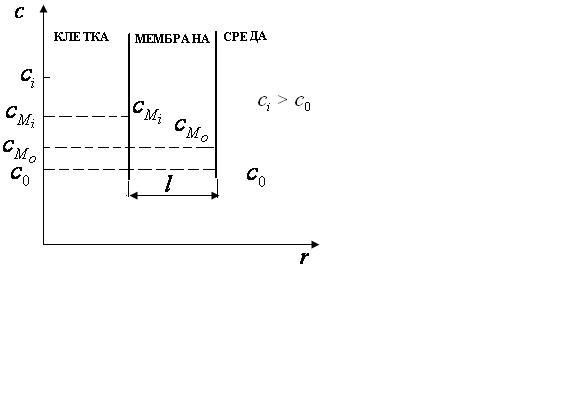 тва
в клетке;
тва
в клетке;
![]() -
концентрация этого же вещества снаружи
клетки;
-
концентрация этого же вещества снаружи
клетки;
![]() -
концентрация этого же вещества внутри
клетки на границе клетка - мембрана;
-
концентрация этого же вещества внутри
клетки на границе клетка - мембрана;
![]() -
концентрация этого же вещества внутри
клетки на границе окружающая среда-мембрана.
-
концентрация этого же вещества внутри
клетки на границе окружающая среда-мембрана.
![]() >
>
![]() .
Следовательно, перенос идет из клетки
в окружающую среду. Для мембраны
.
Следовательно, перенос идет из клетки
в окружающую среду. Для мембраны
![]() .
.
Измерить
![]() и
и![]() трудно,
но зато экспериментально можно измерить
трудно,
но зато экспериментально можно измерить![]() и
и![]() .
Кроме того, считают, что
.
Кроме того, считают, что
![]()
![]() -
коэффициент распределения частиц между
мембраной и окружающей средой.
Следовательно,
-
коэффициент распределения частиц между
мембраной и окружающей средой.
Следовательно,
![]()
Тогда
![]() .
.
Обозначим
![]() - проницаемость мембраны, получим
- проницаемость мембраны, получим
![]()
- это и есть уравнение Коллендора-Берлунда.
Перенос заряженных частиц (ионов) через мембрану
Проникновение заряженных частиц через мембрану (а это ионы), через мембрану зависит не только от концентрационного градиента, но и от градиента электрического потенциала. Плотность потока вещества при этом следует рассчитывать как сумму двух слагаемых:
![]()
Здесь
![]() - плотность потока вещества, обусловленная
градиентом концентраций.
- плотность потока вещества, обусловленная
градиентом концентраций.![]() - плотность потока вещества, обусловленная
градиентом электрического потенциала.
- плотность потока вещества, обусловленная
градиентом электрического потенциала.
Откуда возникает на мембране
электрический градиент, т.е. разность
потенциалов? Согласно полиэлектролитной
теории основой цитоплазмы является
комплексный полиэлектролитный гель
сетчатой структуры с фиксированными
на ней отрицательными зарядами, который
способен избирательно накаплиаить ионы
![]() .
В результате, на наружней поверхности
мембраны скапливается положительный
заряд и положительный потенциал, а на
внутренней - отрицательный потенциал
(позже мы покажем это доказательно).Поэтому вокруг мембраны возникает
.
В результате, на наружней поверхности
мембраны скапливается положительный
заряд и положительный потенциал, а на
внутренней - отрицательный потенциал
(позже мы покажем это доказательно).Поэтому вокруг мембраны возникает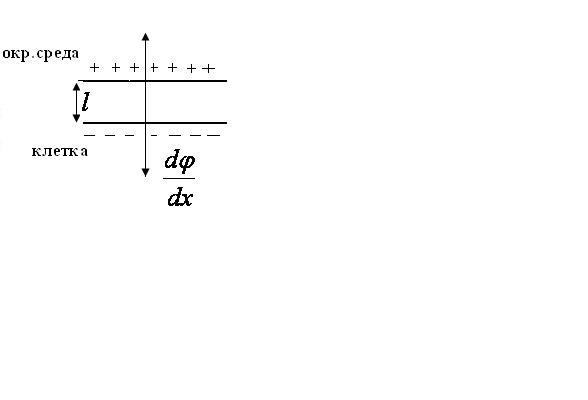
электричкское поле напряженностью
![]() .
Это поле убывет по силе при удалении от
мембраны. Между напряженностью поля и
градиентом потенциала имеется связь:
.
Это поле убывет по силе при удалении от
мембраны. Между напряженностью поля и
градиентом потенциала имеется связь:![]() .
.
Это электрическое поле действует на
ионы с силой
![]() ,
ускоряя или замедляя их (
,
ускоряя или замедляя их (![]() заряд всех ионов).
заряд всех ионов).
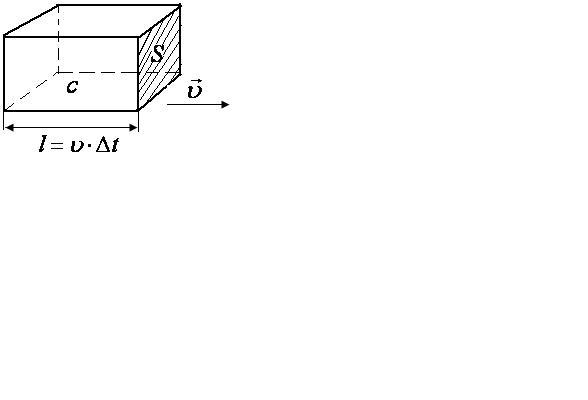
Возьмем 1 моль ионов, в котором содержится
![]() ионов. Чтобы найти поток ионов, выделим
обеъм электролита в виде прямоугольного
параллепипеда с ребом
ионов. Чтобы найти поток ионов, выделим
обеъм электролита в виде прямоугольного
параллепипеда с ребом![]() (
(![]() - скорость движения ионов) и площадью
основания
- скорость движения ионов) и площадью
основания![]() .
Пусть за время
.
Пусть за время![]() ,
все ионы, находящиеся в этом объёме,
пройдут через площадку
,
все ионы, находящиеся в этом объёме,
пройдут через площадку![]() .
Будем считать, что концентрация ионов
равна
.
Будем считать, что концентрация ионов
равна![]() ,
следовательно масса 1 моля ионов
определится как
,
следовательно масса 1 моля ионов
определится как
![]() ,
,
а поток ионов как
![]() .
Плотность потока ионов тогда будет
.
Плотность потока ионов тогда будет
![]() .
.
Скорость
![]() направленного движения ионов
пропорциональна действующей силе
направленного движения ионов
пропорциональна действующей силе![]() ,
где
,
где![]() - сила, действующая на 1 ион,
- сила, действующая на 1 ион,![]() заряд иона.
заряд иона.
![]() ,
,
где
![]() - подвижность ионов.
- подвижность ионов.
![]() ,
где
,
где
![]() -
зарядовое число иона,
-
зарядовое число иона,![]() - заряд электрона. Тогда
- заряд электрона. Тогда
![]() .
.
Здесь![]() -числоФарадея
(заряд 1 моля ионов). Следовательно,
-числоФарадея
(заряд 1 моля ионов). Следовательно,
![]() ,
а
,
а
![]() .
.
Это уравнение называется уравнением Нернста-Планка.
Совокупность концентрационного и электрического градиентов называется градиентом электрохимического потенциала.
Виды диффузии.
1. Простая диффузия – диффузия, при которой молекулы
диффундирующего вещества движутся
без образования комплекса с другими молекулами. В живой клетке такая диффузия обеспечивает прохождение кислорода, лекарственных веществ и ядов через мембрану. Механизм такой диффузии простой: в жидкой фазе молекулы фосфолипидов могут образовывать полости (кинки), в которые способны внедряться молекулы перечисленных веществ. Эти кинки движутся поперек мембраны и переносят диффундирующее вещество. Простая диффузия протекает медленно и не может в достаточном количестве обеспечить клетку питательными веществами. Но природа обеспечила другие виды диффузии.
Диффузия через каналы.

Наличие каналов увеличивает проницаемость мембран.
Облегченная диффузия.
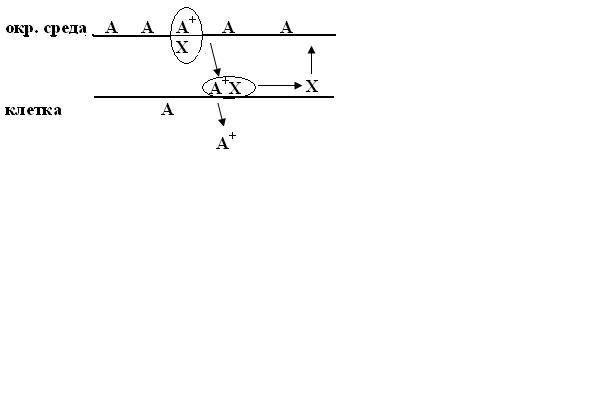
Вещество А самостоятельно слабо диффундирует через мембрану. Но скорость диффузии значительно возрастает, когда молекула А+ этого вещества образует комплекс с некоторым вспомогательным веществом Х, которое растворено в липиде. Этот комплекс диффундирует в мембрану, достигает её противоположной стороны, здесь молекула А освобождается и выходит в клетку, а молекула Х освободившись, диффундирует обратно к наружней стороне мембраны, где снова вступает во взаимодействие с другой молекулой А и процесс повторяется.
Переносчики Х могут быть фиксированными и образовывать пору:
Молекула А+ захватывается
ближайшей 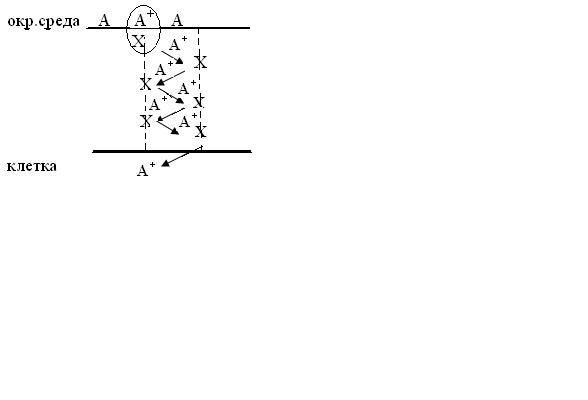 молекулой
Х и переходит внутри мембраны от одной
молекулы Х к другой по эстафете, достигает
противоположной стороны мембраны, где
выходит в клетку.
молекулой
Х и переходит внутри мембраны от одной
молекулы Х к другой по эстафете, достигает
противоположной стороны мембраны, где
выходит в клетку.
Размеры поры не должны превышать размеров молекулы А.
Молекулы переносчики называются ионофорами.
Обменная диффузия.
Вспомогательное вещество Х образует
комплекс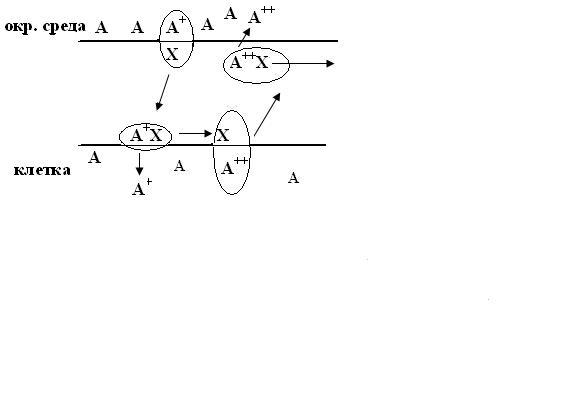 с
молекулой проникающего вещества А+,
комплекс диффундирует через мембрану.
На другой стороне мембраны молекула
А+, освободившись,уходит
в клетку, а ионофор берёт из клетки
другую молекулу А++ и
переносит её в окружающую среду. В
результате концентрация вещества А по
обе стороны мембраны не меняется,
следовательно, этот вид диффузии,
фактически не принимает участия в обмене
веществ.
с
молекулой проникающего вещества А+,
комплекс диффундирует через мембрану.
На другой стороне мембраны молекула
А+, освободившись,уходит
в клетку, а ионофор берёт из клетки
другую молекулу А++ и
переносит её в окружающую среду. В
результате концентрация вещества А по
обе стороны мембраны не меняется,
следовательно, этот вид диффузии,
фактически не принимает участия в обмене
веществ.
Все рассмотренные виды диффузии описываются уравнением Фика, если переносится молекулы или атомы, и уравнением Нернста-Планка, если переносятся ионы.
5.Осмос –движение молекул воды (растворителя) через полупроницаемую мембрану из области большей концентрации растворённого вещества в область меньшей концентрации растворенного вещества. Т.е. это тоже диффузия, но диффузия растворителя.
Сила, которая вызывает это движение растворителя, называется осмотическим давлением.
Плотность потока вещества определяется как
![]() .
.
Здесь
![]() -
коэффициент проницаемости;
-
коэффициент проницаемости;![]() и
и![]() - осмотическое давление по одну и другую
сторону мембраны, соответственно.
- осмотическое давление по одну и другую
сторону мембраны, соответственно.
6. Фильтрация –движение молекул воды (растворителя) через полупроницаемую мембрану из области большей концентрации растворённого вещества в область меньшей концентрации растворенного вещества при наличии гидростатического давления (давления, обусловленного столбом жидкости).
формулу
Явления фильтрации и осмоса имеют особое значение в процессе обмена водой между кровью и тканью.
Активный транспорт.
Пассивный транспорт веществ всегда
стремится выровнять неравномерность
в распределении вещества между клеткой
и средой. Но клеточное содержимое резко
отличается по своему составу от окружающей
клетку среды. В клетке в большом количестве
находятися ионы
![]() ,
недиффундирующие ионы белков, фосфолипидов,
анионы аминокислот и др, содержание
которых в жидкости, окружающей клетку,
незначительно. Другие вещества, наоборот,
в значительно более высоких концентрациях
содержатся в окружающей клетку жидкости,
например,
,
недиффундирующие ионы белков, фосфолипидов,
анионы аминокислот и др, содержание
которых в жидкости, окружающей клетку,
незначительно. Другие вещества, наоборот,
в значительно более высоких концентрациях
содержатся в окружающей клетку жидкости,
например,![]() .
.
В результате такого неравномерного распределения концентраций ионов между клеткой и окружающей средой пассивный перенос не может полностью обеспечить равномерное распределение концентраций. Поэтому в организме одновременно с пассивным транспортом происходит активный транспорт.
Активный транспорт обеспечивает перенос молекул и ионов из области меньших концентраций и электрических потенциалов в область больших концентраций и электрических потенциалов.
Для осуществления такого транспорта клетка совершает работу против градиентов концентраций и потенциалов.
Если через клетку переносится незаряженная частица (атом или молекула), то эта работа равна
![]() ,
,
где
![]() -
количество молей вещества, перенесенного
через мембрану из области меньших
концентраций
-
количество молей вещества, перенесенного
через мембрану из области меньших
концентраций![]() в область больших концентраций
в область больших концентраций![]() ;
;![]() - универсальная газовая постоянная,
- универсальная газовая постоянная,![]() - абсолютная температура.
- абсолютная температура.
Если переносится ион через электрически заряженную мембрану, то эта работа равна
![]() ,
,
где
![]() - валентность ионов,
- валентность ионов,![]() - число Фарадея (заряд 1 моля ионов),
- число Фарадея (заряд 1 моля ионов),![]() разность потенциало между поверхностыми
мембраны.
разность потенциало между поверхностыми
мембраны.![]() зависит от знака заряда ионов.
зависит от знака заряда ионов.
Чтобы совершить эту работу, клетке
нужна энергия. Эту энергию клетка
получает пригидролизе фермента
![]() (адезинтрифосфатоза).
(адезинтрифосфатоза).
Особое внимание следует обратить на
активный транспорт ионов
![]() и
и![]() ,
калий – натриевый насос, т.к. именно эти
ионы играют большую роль при генерации
биоэлектрических потенциалов и проведении
возбуждения.
,
калий – натриевый насос, т.к. именно эти
ионы играют большую роль при генерации
биоэлектрических потенциалов и проведении
возбуждения.
Переход
![]() из клетки зависит от концентрации
из клетки зависит от концентрации![]() во внешней среде, а переход
во внешней среде, а переход![]() в клетку, в свою очередь, эависит от
концентрации
в клетку, в свою очередь, эависит от
концентрации![]() в цитоплазме.
в цитоплазме.
Предполагают, что перенос
![]() и
и![]() осуществляется специальным переносчиком
белковой или белково-липидной природы.
осуществляется специальным переносчиком
белковой или белково-липидной природы.
Рассмотрим перенос
![]() из окружающей среды в клетку. Он начинается
на внутренней поверхности мембраны и
происходит в три стадии.
из окружающей среды в клетку. Он начинается
на внутренней поверхности мембраны и
происходит в три стадии.
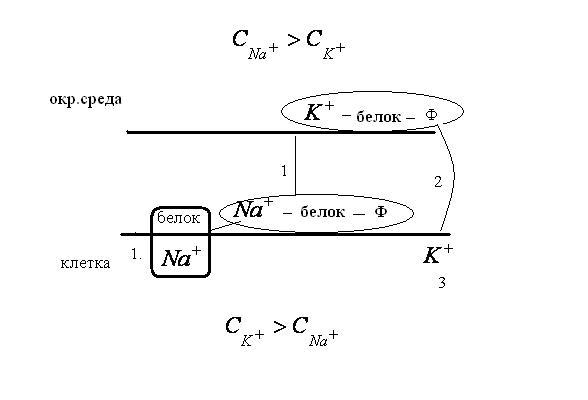
Киназная. Переносчик на внутренней
стороне мембраны захватывет из цитоплазмы
ион
![]() :
:
![]()
Комплекс
![]() переносится на наружнюю поверхность
мембраны за счет гидролиза
переносится на наружнюю поверхность
мембраны за счет гидролиза![]() .
.
Ионообменная. На наружней поверхности
мембраны ионы
![]() обмениваются на ионы
обмениваются на ионы![]()
![]()
Комплекс
![]() снова движется к внутренней стороне
мембраны.
снова движется к внутренней стороне
мембраны.
3. Фосфатозная. Эта фаза заканчивает
цикл на внутренней поверхности мембраны
дефосфолированием переносчика и
освобождением ионов
![]()
![]()
Лекция 8
Электромагнитные явления в биологических системах.
Природа биопотенциалов и способы их описания.
Все процессы жизнедеятельности организмов сопровождаются появлением в клетках и тканях электродвижущих сил. Электрические явления играют большую роль в важнейших физиологических процессах: возбуждение клеток и проведение возбуждения по клеткам.
В возникновении биопотенциалов решающую роль играет разность потенциалов, обусловленная несимметричным распределением ионов. К таким разностям потенциалов относятся диффузионные, мембранные и фазовые.
1. Диффузионные потенциалывозникают
на границе раздела двух жидких сред в
результате различной подвижности ионов.
Рассмотрим пример: Имеется раствор
серной кислоты, разделенный пористой
перегородкой. Пусть концентрация![]() в левой части больше, чем в прав
в левой части больше, чем в прав ой.
Ионы
ой.
Ионы![]() и
и![]() будут диффундировать из левой части в
правую часть сосуда с разной скоростью
по градиенту концентрации. Скорость
диффузии определяется подвижностью
ионов. Подвижность ионов
будут диффундировать из левой части в
правую часть сосуда с разной скоростью
по градиенту концентрации. Скорость
диффузии определяется подвижностью
ионов. Подвижность ионов![]() больше подвижности ионов
больше подвижности ионов![]() :
:![]() Следовательно, ионы
Следовательно, ионы![]() будут намного опережать ионы
будут намного опережать ионы![]() .
По обе стороны перегородки установятся
потенциалы: слева “-“ справа “+”.
Возникает диффузионная разность
потенциалов. Эта разность потенциалов
будет замедлять “быстрые“ ионы и
ускорять “медленные“, т.е. возникающее
электрическое поле направлено против
сил диффузии. Диффузионная разность
потенциалов максимален в тот момент,
когда скорости диффузии становятся
равными:
.
По обе стороны перегородки установятся
потенциалы: слева “-“ справа “+”.
Возникает диффузионная разность
потенциалов. Эта разность потенциалов
будет замедлять “быстрые“ ионы и
ускорять “медленные“, т.е. возникающее
электрическое поле направлено против
сил диффузии. Диффузионная разность
потенциалов максимален в тот момент,
когда скорости диффузии становятся
равными:
![]() .
.
Здесь
![]() - ;подвижность катионов;
- ;подвижность катионов;![]() - подвижность анионов;
- подвижность анионов;![]() - универсальная газовая постоянная;
- универсальная газовая постоянная;![]() - абсолютная температура;
- абсолютная температура;![]() -валент-ность ионов;
-валент-ность ионов;![]() -
число Фарадея;
-
число Фарадея;![]() - активная концентрация в области откуда
идет диффузия;
- активная концентрация в области откуда
идет диффузия;![]() - активная концентрация в области куда
идет диффузия.
- активная концентрация в области куда
идет диффузия.
2. Мембранный потенциал. В рассматриваемом
примере пористую перегородку заменим
полупроницаемой мембраной, пропускающую
только катионы – положительно заряженные
ионы (это может быть мембрана с большой
концентрацией фиксированных отрицательных
ионов).
В этом случае подвижность ионов при
переходе через мембрану не является
определяющим фактором – в правую часть
переходят только положительно заряженные
ионы водорода. Через некоторое время
диффузия ионов
![]() прекратится, т.к. они испытывают притяжение
со стороны оставшихся в левой части
ионов
прекратится, т.к. они испытывают притяжение
со стороны оставшихся в левой части
ионов![]() .
Поэтому установится равновесие , в
результате чего возникает двойной
электрический слой: слева заряды “-“,
справа заряды “+”, между сторонами
мембраны возникнет разность потенциалов,
называемая мембранным потенциалом,
величина которого есть
.
Поэтому установится равновесие , в
результате чего возникает двойной
электрический слой: слева заряды “-“,
справа заряды “+”, между сторонами
мембраны возникнет разность потенциалов,
называемая мембранным потенциалом,
величина которого есть
![]() уравнение Нернста.
уравнение Нернста.
3. Фазовый потенциал возникает на границе раздела двух несмешивающихся фаз (например, раствор электролита в воде и какое-нибудь масло) в результате различной растворимости анионов и катионов в неводной фазе. Если, например, катионы растворимы в неводной фазе, то они активнее перейдут в нее и зарядят ее положительно относительно водной фазы. Наблюдается фазовый потенциал при механическом повреждении мембраны.
Равенство Доннана.
При выводе уравнений, описывающих
распределение ионов между клеткой и
окружающей средой, выполняется условие
электронейтральности, равенство
суммарной концентрации анионов ( в
основном
![]() и ионов макромолекул
и ионов макромолекул![]() )
катионов
)
катионов![]()
как внутри клетки
![]() ,
,
так и снаружи клетки
![]() .
.
Здесь
![]()
![]() --
число отрицательных зарядов на каждой
белковой молекуле.
--
число отрицательных зарядов на каждой
белковой молекуле.
В межклеточной жидкости содержание
катионов значительно выше, чем ионов
макромолекул
![]() .
Поэтому
.
Поэтому
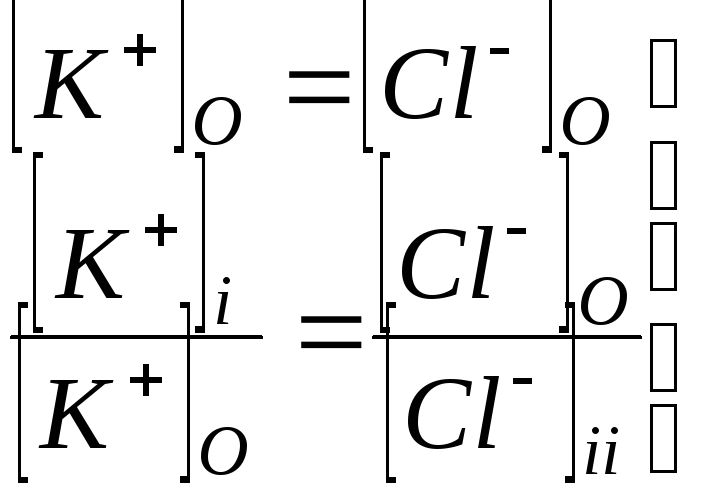 - равенство Доннана
- равенство Доннана
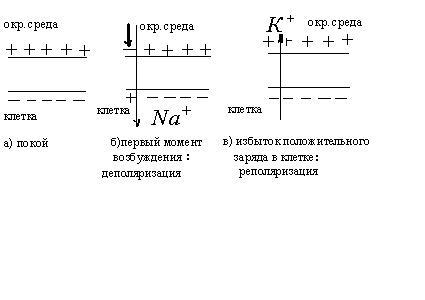
Потенциал покоя
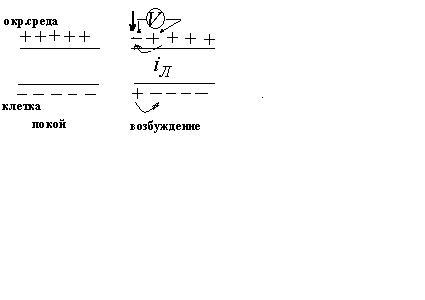
Экспериментально установлено, что цитоплазма в состоянии покоя имеет отрицательный потенциал, а окружающая среда- положительный.
Действительно, в первом приближении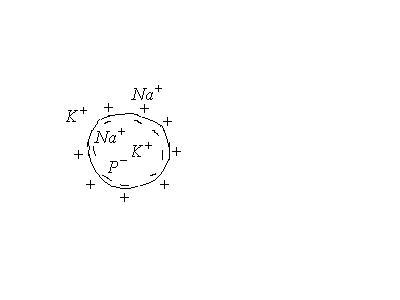
В клетке
![]() в 20-40 раз.
в 20-40 раз.
Снаружи
![]() в 10 раз.
в 10 раз.
Это неравномерное распределение
концентраций обусловлено
![]() насосом, при котором при переносе
насосом, при котором при переносе![]() переносится
переносится![]() .
Кроме того, в клетке имеются анионы
макромолекул
.
Кроме того, в клетке имеются анионы
макромолекул![]() (белков, аминокислот, и др.).
(белков, аминокислот, и др.).
Ионы
![]() внутри клетки не связаны с другими
ионами и могут диффундировать туда, где
их мало, т.е. в окружающую среду. Т.е. в
состоянии покоя клетка проницаема
только для ионов
внутри клетки не связаны с другими
ионами и могут диффундировать туда, где
их мало, т.е. в окружающую среду. Т.е. в
состоянии покоя клетка проницаема
только для ионов![]() .
Анионы не могут проникать через мембрану
и остаются на внутренней поверхности
мембраны. Т.о. мембрана снаружи зарядится
положительно, а внутри – отрицательно.
.
Анионы не могут проникать через мембрану
и остаются на внутренней поверхности
мембраны. Т.о. мембрана снаружи зарядится
положительно, а внутри – отрицательно.
Между внутренней и внешней поверхностями
мембраны возникает разность потенциалов
мембранной природы. Эта разность
потенциалов между клеткой и окружающей
средой, измеренная в состоянии
физиологического покоя, называется
потенциалом покоя.
Если принять, в первом приближении, что
потенциал покоя определяется только
диффузией ионов
![]() ,
то величина потенциала покоя определяется
как
,
то величина потенциала покоя определяется
как
 уравнение
Нернста.
уравнение
Нернста.
![]() - активная концентрация ионов
- активная концентрация ионов![]() внутри клетки,
внутри клетки,
![]() - активная концентрация ионов
- активная концентрация ионов![]() снаружи.
снаружи.
Если
![]() =
=![]() ,
то
,
то![]() клетка мертва.
клетка мертва.
Но
![]() ,
поэтому
,
поэтому![]() .
Например, для аксона гигантского кальмара
.
Например, для аксона гигантского кальмара![]()
![]() ,
а это подтверждает, что в основе
возникновения потенциала действия
лежит перенос ионов.
,
а это подтверждает, что в основе
возникновения потенциала действия
лежит перенос ионов.
В реальности в состоянии покоя мембрана
проницаема не только для ионов
![]() ,
но и для ионов
,
но и для ионов![]() и
и![]() .
Например, для аксона гигантского кальмара
экспериментально установлено, что
.
Например, для аксона гигантского кальмара
экспериментально установлено, что![]() .
Основной вклад в потенциал покоя вносят
ионы
.
Основной вклад в потенциал покоя вносят
ионы![]() и
и![]() .
Ионов
.
Ионов![]() переносится очень мало. Поэтому на
переносится очень мало. Поэтому на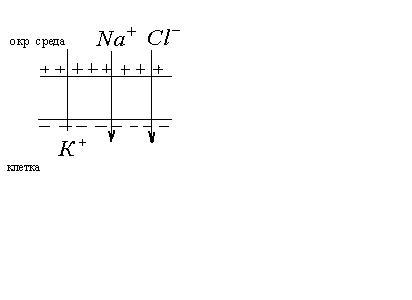 внешней
поверхности сосредоточивается
положительный заряд, а на внутренней –
отрицательный. Потенциал покоя
определяется тремя диффузионными
потоками и вычисляется (с учетом равенства
Доннана) по формуле
внешней
поверхности сосредоточивается
положительный заряд, а на внутренней –
отрицательный. Потенциал покоя
определяется тремя диффузионными
потоками и вычисляется (с учетом равенства
Доннана) по формуле
 .
.
Кроме простой диффузии ионов
![]() и
и![]() может
идти обменная диффузия, но, как мы
отмечали, потоки их равны и обменная
диффузия не влияет на мембранный
потенциал.
может
идти обменная диффузия, но, как мы
отмечали, потоки их равны и обменная
диффузия не влияет на мембранный
потенциал.
Потенциал действия.
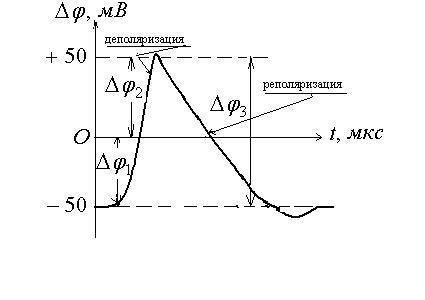
Все клетки возбудимых тканей (нервная, мышечная, железистая) под действием различных раздражителей достаточной силы способны переходить в возбужденное состояние. Обязательным признаком возбуждения является изменение электрического состояния мембраны.
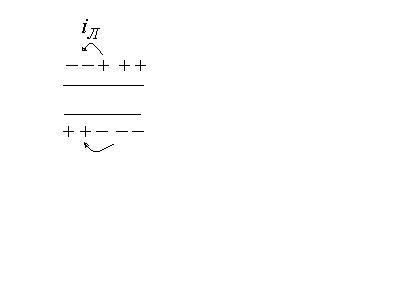
Опыт показывает, что возбужденный
участок становится электроотри-цательным
по отношению к невозбужденному участку.
Следовательно, на возбужденном участке
происходит перераспределение ионов.
При возбуждении  это
перераспределение кратковременно и
концентрации восстанавливаются после
снятия возбуждения, а разность потенциалов
становится равной исходной, т.е. потенциалу
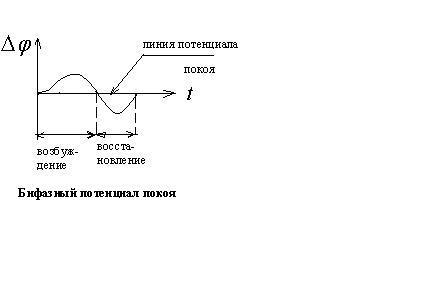
покоя. Для аксона кальмара обнаружена
такая зависимость изменения потенциала
при возбуждении от времени
это
перераспределение кратковременно и
концентрации восстанавливаются после
снятия возбуждения, а разность потенциалов
становится равной исходной, т.е. потенциалу
покоя. Для аксона кальмара обнаружена
такая зависимость изменения потенциала
при возбуждении от времени
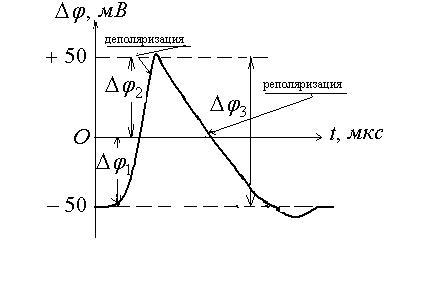
![]() - потенциал покоя;
- потенциал покоя;![]() мембранный потенциал при возбуждении;
мембранный потенциал при возбуждении;
![]()
![]() - общее изменение разности потенциалов.
- общее изменение разности потенциалов.
Общее изменение разности потенциалов между клеткой и окружающей средой, происходящее при пороговом и сверхпороговом возбуждении клетки, называется потенциалом действия.
Механизм возникновения потенциала действия. В 1938 году Круэл и Картис показали, что сопротивление аксона кальмара в состоянии покоя 1000 Ом/см2, а при возбуждении 25 Ом/см2, т.е. уменьшается в 40 раз. При этом сопротивление цитоплазмы не изменяется. Следовательно, уменьшение сопротивления мембраны обусловлено только её проницаемости для ионов, т.к. именно они являются переносчиками электричества в мембранах и клетках.
Хаджкин, Хаксли и Катц показали, что при
возбуждении про-ницаемость мембраны
увеличивается только для ионов
![]() ,
причем в 500 раз. Это приводит к увеличению
диффузии ионов
,
причем в 500 раз. Это приводит к увеличению
диффузии ионов![]() из
окружающей среды в клетку (по
концентрационному градиенту), что
приводит к изменению потенциала мембраны.
В первые моменты возбуждения интенсивность
потока ионов
из
окружающей среды в клетку (по
концентрационному градиенту), что
приводит к изменению потенциала мембраны.
В первые моменты возбуждения интенсивность
потока ионов![]() из клетки остается такой же, как и до
возбуждения.
из клетки остается такой же, как и до
возбуждения.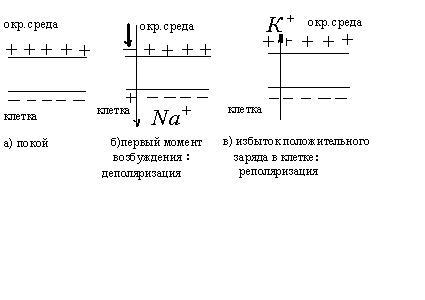
Поэтому поток ионов
![]() вызывает
исчезновение избыточного отрицательного
потенциала на внутренней поверхности
мембраны. Эта фаза называется деполяризацией
и длится короткое время. Затем начинается
другая фаза – реполяризация., заключающаяся
в следующем. Диффузия ионов
вызывает
исчезновение избыточного отрицательного
потенциала на внутренней поверхности
мембраны. Эта фаза называется деполяризацией
и длится короткое время. Затем начинается
другая фаза – реполяризация., заключающаяся
в следующем. Диффузия ионов![]() внутрь клетки нарушает равновесие
концентраций в клетке. В связи с этим
повышается проницаемость мембраны для
ионов
внутрь клетки нарушает равновесие
концентраций в клетке. В связи с этим
повышается проницаемость мембраны для
ионов![]() ,
начинается диффузия ионов
,
начинается диффузия ионов![]() из клетки в окружающую среду. Поток
ионов
из клетки в окружающую среду. Поток
ионов![]() из клетки приводит к уменьшению
проницаемости для ионов
из клетки приводит к уменьшению
проницаемости для ионов![]() .
В результате происходит реполяризация
мембраны и восстановление потенциала
покоя. Проницаемость мембраны для ионов
.
В результате происходит реполяризация
мембраны и восстановление потенциала
покоя. Проницаемость мембраны для ионов![]() и
и![]() падает до исходной величины. Фаза
реполяризации длится дольше фазы
деполяризации, поэтому и кривая более
пологая.
падает до исходной величины. Фаза
реполяризации длится дольше фазы
деполяризации, поэтому и кривая более
пологая.
В некоторых случаях регистрируется так
называемый следовой потенциал, как на
данном рисунке. Он вызван тем, что после
окончания возбуждения проницаемость
мембраны для ионов
![]() и
и![]() остпется повышенной.
остпется повышенной.
Т.о. формирование потенциала действия
обусловлено двумя потоками через
мембрану: поток
![]() внутрь клетки приводит к перезарядке
мембраны, а противоположный поток
внутрь клетки приводит к перезарядке
мембраны, а противоположный поток![]() обусловливает восстановление потенциала.
Потоки эти приблизительно равны по
величине, но сдвинуты по времени.
Благодаря этому сдвигу во времени и
возможно появление потенциала действия.
обусловливает восстановление потенциала.
Потоки эти приблизительно равны по
величине, но сдвинуты по времени.
Благодаря этому сдвигу во времени и
возможно появление потенциала действия.
Распространение потенциала действия.
(проведение возбуждения по нервным волокнам).
Потенциал действия, возникнув в одном
участке нервной клетки, распространяется
по всей её поверхности.
В результате возбуждения между
возбужденным и невозбужденным участками
возникает разность потенциалов. Эта
разность потенциалов создает электрический
ток, называемый локальным током (от
невозбужденного участка к возбужденному).
Локальный ток оказывает на соседний
невозбужденный участок такое же действие
как и исходный возбудитель и увеличивает
проница-емость мембраны для ионов
![]() .
В результате и в этом участке снижкется
потенциал покоя и возникает потенциал
действия.
.
В результате и в этом участке снижкется
потенциал покоя и возникает потенциал
действия.
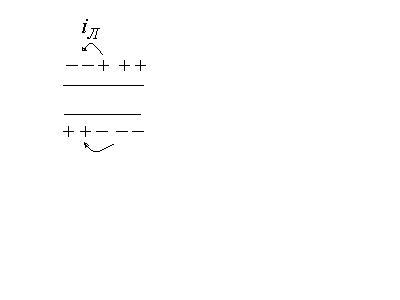
В участке, который был ранее возбужден, происходят восстановительные процессы реполяризации.
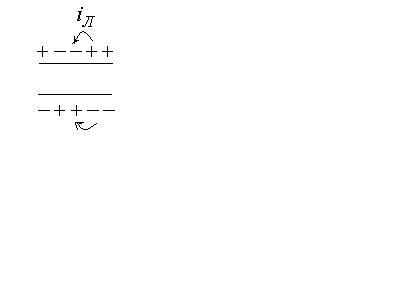
Э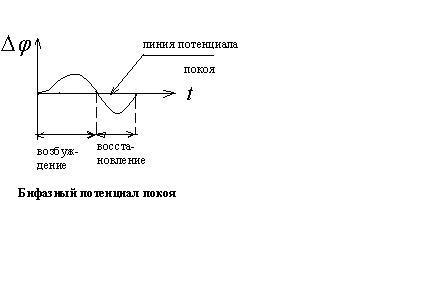 тот
процесс повторяется многократно и
обусловливает распространение импульсов
по всей длине клетки в обоих направлениях.
Разность потенциалов между возбужденным
и невозбужденным участками изменяется
как
тот
процесс повторяется многократно и
обусловливает распространение импульсов
по всей длине клетки в обоих направлениях.
Разность потенциалов между возбужденным
и невозбужденным участками изменяется
как
Волна возбуждения под влиянием локальных токов распространяется по нервному волокну без затухания. Это обусловлено тем, что локальные токи только деполяризуют мембрану, а потенциал действия в каждом участке мембраны поддерживается независимыми потоками, перпендикулярно направлению распространения возбуждения, т.е. источником энергии такой электромагнитной волны является сама среда.
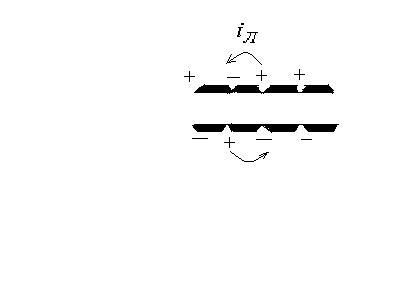
Если в нервных волокнах нет миелиновых оболочек, то возбуждение в них распространяется так, как мы рассмотрели.
Если же имеются миелиновые оболочки (миелин – жироподобное вещество - диэлектрик), то локальные токи распространяются между участками, на которых оболочки прерываются– между перехватами Ранвье, т.к. миелин является изолятором:
Лекция 8
Электромагнитные явления в биологических системах
Природа биопотенциалов и способы их описания
Все процессы жизнедеятельности организмов сопровождаются появлением в клетках и тканях электродвижущих сил. Электрические явления играют большую роль в важнейших физиологических процессах: возбуждение клеток и проведение возбуждения по клеткам.
В возникновении биопотенциалов решающую роль играет разность потенциалов, обусловленная несимметричным распределением ионов. К таким разностям потенциалов относятся диффузионные, мембранные и фазовые.
1. Диффузионные потенциалывозникают на границе раз-
дела двух жидких сред в результате
различной подвижности ионов. Рассмотрим
пример: имеется раствор серной кислоты,
разделенный пористой перегородкой.
Пусть концентрация
![]() в левой части больше, чем в правой. Ионы
в левой части больше, чем в правой. Ионы![]() и
и![]() будут диффундироватьи
будут диффундироватьи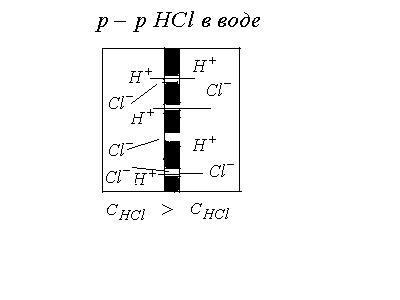 з
левой части в правую часть сосуда с
разной скоростью по градиенту концентрации.
Скорость диффузии определяется
подвижностью ионов. Подвижность ионов
з
левой части в правую часть сосуда с
разной скоростью по градиенту концентрации.
Скорость диффузии определяется
подвижностью ионов. Подвижность ионов![]() больше подвижности ионов
больше подвижности ионов![]() :
:![]() Следовательно, ионы
Следовательно, ионы![]() будут намного опережать ионы
будут намного опережать ионы![]() .
По обе стороны перегородки установятся
потенциалы: слева “-“ справа “+”.
Возникает диффузионная разность
потенциалов. Эта разность потенциалов
будет замедлять “быстрые“ ионы и
ускорять “медленные“, т.е. возникающее
электрическое поле направлено против
сил диффузии. Диффузионная разность
потенциалов максимален в тот момент,
когда скорости диффузии становятся
равными:
.
По обе стороны перегородки установятся
потенциалы: слева “-“ справа “+”.
Возникает диффузионная разность
потенциалов. Эта разность потенциалов
будет замедлять “быстрые“ ионы и
ускорять “медленные“, т.е. возникающее
электрическое поле направлено против
сил диффузии. Диффузионная разность
потенциалов максимален в тот момент,
когда скорости диффузии становятся
равными:
![]() .
.
Здесь
![]() - ;подвижность катионов;
- ;подвижность катионов;![]() - подвижность анионов;
- подвижность анионов;![]() - универсальная газовая постоянная;
- универсальная газовая постоянная;![]() - абсолютная температура;
- абсолютная температура;![]() -валентность ионов;
-валентность ионов;![]() -
число Фарадея;
-
число Фарадея;![]() - активная концентрация в области откуда
идет диффузия;
- активная концентрация в области откуда
идет диффузия;![]() - активная концентрация в области куда
идет диффузия.
- активная концентрация в области куда
идет диффузия.
2. Мембранный потенциал. В рассматриваемом примере пористую перегородку заменим полупроницаемой мембраной, пропускающую только катионы – положительно заряженные ионы (это может быть мембрана с большой концентрацией фиксированных отрицательных ионов).
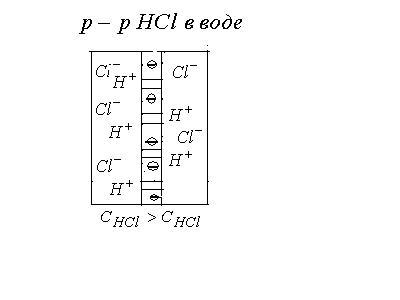
В этом случае подвижность ионов при
переходе через мембрану не является
определяющим фактором – в правую часть
переходят только положительно заряженные
ионы водорода. Через некоторое время
диффузия ионов
![]() прекратится, т.к. они испытывают притяжение
со стороны оставшихся в левой части
ионов
прекратится, т.к. они испытывают притяжение
со стороны оставшихся в левой части
ионов![]() .
Поэтому установится равновесие , в
результате чего возникает двойной
электрический слой: слева заряды “-“,
справа заряды “+”, между сторонами
мембраны возникнет разность потенциалов,
называемая мембранным потенциалом,
величина которого есть
.
Поэтому установится равновесие , в
результате чего возникает двойной
электрический слой: слева заряды “-“,
справа заряды “+”, между сторонами
мембраны возникнет разность потенциалов,
называемая мембранным потенциалом,
величина которого есть
![]() уравнение Нернста.
уравнение Нернста.
3. Фазовый потенциал возникает на границе раздела двух несмешивающихся фаз (например, раствор электролита в воде и какое-нибудь масло) в результате различной растворимости анионов и катионов в неводной фазе. Если, например, катионы растворимы в неводной фазе, то они активнее перейдут в нее и зарядят ее положительно относительно водной фазы. Наблюдается фазовый потенциал при механическом повреждении мембраны.
Равенство Доннана.
При выводе уравнений, описывающих
распределение ионов между клеткой и
окружающей средой, выполняется условие
электронейтральности, равенство
суммарной концентрации анионов ( в
основном
![]() и ионов макромолекул
и ионов макромолекул![]() )
катионов
)
катионов![]()
как внутри клетки
![]() ,
,
так и снаружи клетки
![]() .
.
Здесь
![]()
![]() -число отрицательных зарядов на каждой
белковой молекуле.
-число отрицательных зарядов на каждой
белковой молекуле.
В межклеточной жидкости содержание
катионов значительно выше, чем ионов
макромолекул
![]() .
Поэтому
.
Поэтому
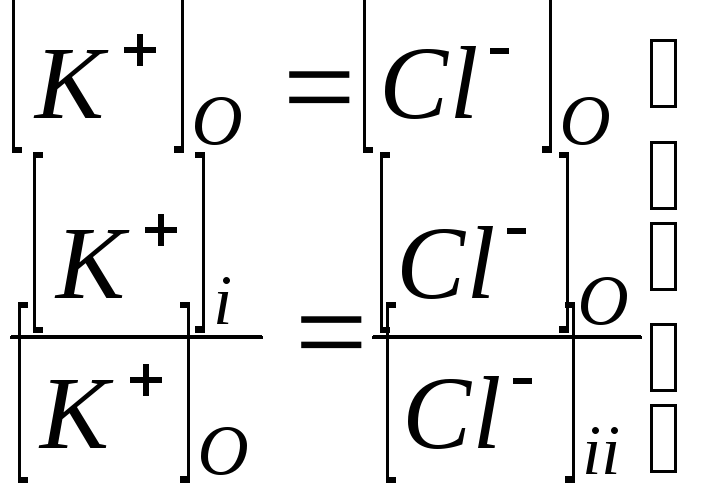 - равенство Доннана
- равенство Доннана
Потенциал покоя
Экспериментально установлено, что цитоплазма в состоянии покоя имеет отрицательный потенциал, а окружающая среда- положительный.
Действительно, в первом приближении
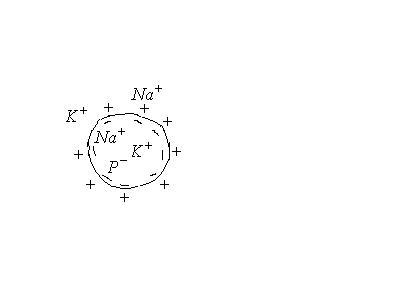
В клетке
![]() в 20-40 раз.
в 20-40 раз.
Снаружи
![]() в 10 раз.
в 10 раз.
Это неравномерное распределение
концентраций обусловлено
![]() насосом, при котором при переносе
насосом, при котором при переносе![]() переносится
переносится![]() .
Кроме того, в клетке имеются анионы
макромолекул
.
Кроме того, в клетке имеются анионы
макромолекул![]() (белков, аминокислот, и др.).
(белков, аминокислот, и др.).
Ионы
![]() внутри клетки не связаны с другими
ионами и могут диффундировать туда, где
их мало, т.е. в окружающую среду. Т.е. в
состоянии покоя клетка проницаема
только для ионов
внутри клетки не связаны с другими
ионами и могут диффундировать туда, где
их мало, т.е. в окружающую среду. Т.е. в
состоянии покоя клетка проницаема
только для ионов![]() .
Анионы не могут проникать через мембрану
и остаются на внутренней поверхности
мембраны. Т.о. мембрана снаружи зарядится
положительно, а внутри – отрицательно.
.
Анионы не могут проникать через мембрану
и остаются на внутренней поверхности
мембраны. Т.о. мембрана снаружи зарядится
положительно, а внутри – отрицательно.
Между внутренней и внешней поверхностями мембраны возникает разность потенциалов мембранной природы. Эта разность потенциалов между клеткой и окружающей средой, измеренная в состоянии физиологического покоя, называется потенциалом покоя.

Если принять, в первом приближении, что
потенциал покоя определяется только
диффузией ионов
![]() ,
то величина потенциала покоя определяется
как
,
то величина потенциала покоя определяется
как
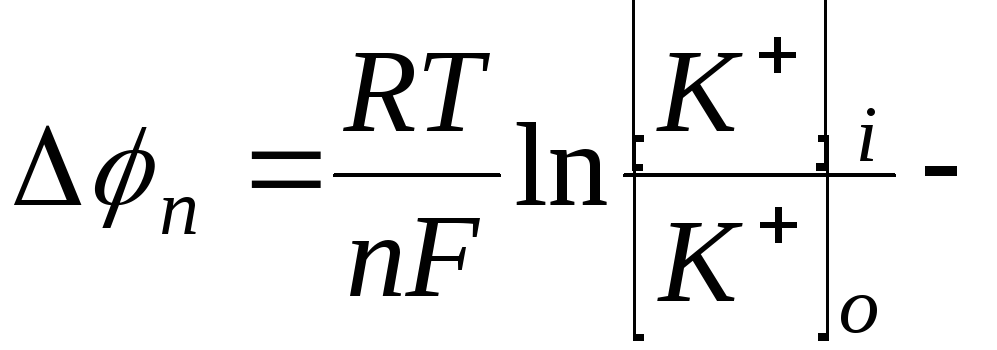 уравнение
Нернста.
уравнение
Нернста.
![]() - активная концентрация ионов
- активная концентрация ионов![]() внутри клетки,
внутри клетки,
![]() - активная концентрация ионов
- активная концентрация ионов![]() снаружи.
снаружи.
Если
![]() =
=![]() ,
то
,
то![]() клетка мертва.
клетка мертва.
Но
![]() ,
поэтому
,
поэтому![]() .
Например, для аксона гигантского кальмара
.
Например, для аксона гигантского кальмара![]()
![]() ,
а это подтверждает, что в основе
возникновения потенциала действия
лежит перенос ионов.
,
а это подтверждает, что в основе
возникновения потенциала действия
лежит перенос ионов.
В реальности в состоянии покоя мембрана
проницаема не только для ионов
![]() ,
но и для ионов
,
но и для ионов![]() и
и![]() .
Например, для аксона гигантского кальмара
экспериментально установлено, что
.
Например, для аксона гигантского кальмара
экспериментально установлено, что![]() .
Основной вклад в потенциал покоя вносят
ионы
.
Основной вклад в потенциал покоя вносят
ионы![]() и
и![]() .
Ионов
.
Ионов![]() переносится очень мало. Поэтому на
внешней поверхности
переносится очень мало. Поэтому на
внешней поверхности
сосредоточивается положительный заряд,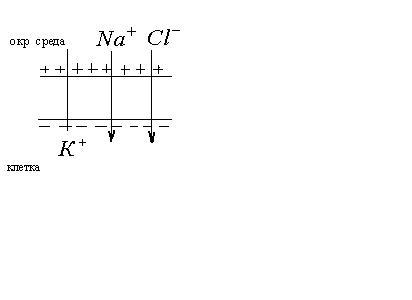 а на внутренней – отрицательный.
Потенциал покоя определяется тремя
диффузионными потоками и вычисляется
(с учетом равенства Доннана) по формуле
а на внутренней – отрицательный.
Потенциал покоя определяется тремя
диффузионными потоками и вычисляется
(с учетом равенства Доннана) по формуле
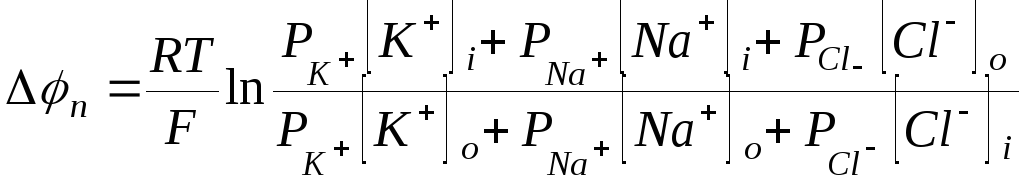 .
.
Кроме простой диффузии ионов
![]() и
и![]() может
идти обменная диффузия, но, как мы
отмечали, потоки их равны и обменная
диффузия не влияет на мембранный
потенциал.
может
идти обменная диффузия, но, как мы
отмечали, потоки их равны и обменная
диффузия не влияет на мембранный
потенциал.
Потенциал действия.
Все клетки возбудимых тканей (нервная, мышечная, железистая) под действием различных раздражителей достаточной силы способны переходить в возбужденное состояние. Обязательным признаком возбуждения является изменение электрического состояния мембраны.
Опыт показывает, что возбужденный
участок становится 
электроотрицательным по отношению к
невозбужденному участку. Следовательно, на возбужденном участке происходит перераспределение ионов. При возбуждении это перераспределение кратковременно и концентрации восстанавливаются после снятия возбуждения, а разность потенциалов становится равной исходной, т.е. потенциалу покоя. Для аксона кальмара обнаружена такая зависимость изменения потенциала при возбуждении от времени
![]() - потенциал покоя;
- потенциал покоя;![]() мембранный потенциал при возбуждении;
мембранный потенциал при возбуждении;
![]()
![]() - общее изменение разности потенциалов.
- общее изменение разности потенциалов.
Общее изменение разности потенциалов между клеткой и окружающей средой, происходящее при пороговом и сверхпороговом возбуждении клетки, называется потенциалом действия.
Механизм возникновения потенциала действия. В 1938 году Круэл и Картис показали, что сопротивление аксона кальмара в состоянии покоя 1000 Ом/см2, а при возбуждении 25 Ом/см2, т.е. уменьшается в 40 раз. При этом сопротивление цитоплазмы не изменяется. Следовательно, уменьшение сопротивления мембраны обусловлено только её проницаемости для ионов, т.к. именно они являются переносчиками электричества в мембранах и клетках.
Хаджкин, Хаксли и Катц показали, что при
возбуждении проницаемость мембраны
увеличивается только для ионов
![]() ,
причем в 500 раз. Это приводит к увеличению
диффузии ионов
,
причем в 500 раз. Это приводит к увеличению
диффузии ионов![]() из
окружающей среды в клетку (по
концентрационному градиенту), что
приводит к изменению потенциала мембраны.
В первые моменты возбуждения интенсивность
потока ионов
из
окружающей среды в клетку (по
концентрационному градиенту), что
приводит к изменению потенциала мембраны.
В первые моменты возбуждения интенсивность
потока ионов![]()
из клетки остается
такой же, как и до возбуждения. Поэтому
поток ионов
![]() вызывает
исчезновение избыточного отрицательного
потенциала на внутренней поверхности
мембраны. Эта фаза называется деполяризацией
и длится короткое время. Затем начинается
другая фаза – реполяризация., заключающаяся
в следующем. Диффузия ионов
вызывает
исчезновение избыточного отрицательного
потенциала на внутренней поверхности
мембраны. Эта фаза называется деполяризацией
и длится короткое время. Затем начинается
другая фаза – реполяризация., заключающаяся
в следующем. Диффузия ионов![]() внутрь клетки нарушает равновесие
концентраций в клетке. В связи с этим
повышается проницаемость мембраны для
ионов
внутрь клетки нарушает равновесие
концентраций в клетке. В связи с этим
повышается проницаемость мембраны для
ионов![]() ,
начинается диффузия ионов
,
начинается диффузия ионов![]() из клетки в окру-жающую среду. Поток
ионов
из клетки в окру-жающую среду. Поток
ионов![]() из клетки приводит к уменьшению
проницаемости для ионов
из клетки приводит к уменьшению
проницаемости для ионов![]() .
В результате происходит реполяризация
мембраны и восстановление потенциала
покоя. Проницаемость мембраны для ионов
.
В результате происходит реполяризация
мембраны и восстановление потенциала
покоя. Проницаемость мембраны для ионов![]() и
и![]() падает до исходной величины. Фаза
реполяризации длится дольше фазы
деполяризации, поэтому и кривая более
пологая.
падает до исходной величины. Фаза
реполяризации длится дольше фазы
деполяризации, поэтому и кривая более
пологая.
В некоторых случаях
регистрируется так называемый следовой
потенциал, как на данном рисунке. Он
вызван тем, что после окончания возбуждения
проницаемость мембраны для ионов
![]() и
и![]() остпется повышенной.
остпется повышенной.
Т.о. формирование
потенциала действия обусловлено двумя
потоками через мембрану: поток
![]() внутрь клетки приводит к перезарядке
мембраны, а противоположный поток
внутрь клетки приводит к перезарядке
мембраны, а противоположный поток![]() обусловливает восстановление потенциала.
Потоки эти приблизительно равны по
величине, но сдвинуты по времени.
Благодаря этому сдвигу во времени и
возможно появление потенциала действия.
обусловливает восстановление потенциала.
Потоки эти приблизительно равны по
величине, но сдвинуты по времени.
Благодаря этому сдвигу во времени и
возможно появление потенциала действия.
Распространение потенциала действия.
(проведение возбуждения по нервным волокнам).
Потенциал действия, возникнув в одном участке нервной клетки, распространяется по всей её поверхности.
В результате
возбуждения между возбужденным и
невозбужденным участками возникает
разность потенциалов. Эта разность
потенциалов создает электрический ток,
называемый локальным током (от
невозбужденного участка к возбужденному).
Локальный ток оказывает на соседний
невозбужденный участок такое же действие
как и исходный возбудитель и увеличивает
проница-емость мембраны для ионов
![]() .
В результате и в этом участке снижкется
потенциал покоя и возникает потенциал
действия.
.
В результате и в этом участке снижкется
потенциал покоя и возникает потенциал
действия.
В участке, который был ранее возбужден, происходят восстановительные процессы реполяризации.

Этот процесс повторяется многократно и обусловливает распространение импульсов по всей длине клетки в обоих направлениях. Разность потенциалов между возбужденным и невозбужденным участками изменяется как
Волна возбуждения под влиянием локальных токов распространяется по нервному волокну без затухания. Это обусловлено тем, что локальные токи только деполяризуют мембрану, а потенциал действия в каждом участке мембраны поддерживается независимыми потоками, перпендикулярно направлению распространения возбуждения, те. Источником энергии такой электромагнитной волны является сама среда.
Если в нервных волокнах нет миелиновых оболочек, то возбуждение в них распространяется так, как мы рассмотрели.
Если же имеются миелиновые оболочки (миелин – жироподобное вещество - диэлектрик), то локальные токи распространяются между участками, на которых оболочки прерываются – между перехватами Ранвье, т.к. миелин является изолятором:

Лекция 9
Биофизические принципы исследования
Электрических полей в организме.
Нервные волокна, мышцы, и, следовательно, клетки, по которым распространяется раздражение, можно моделировать как электрический диполь. Действительно:

Электрический диполь
Электрический диполь- система двух точечных зарядов, равных по величине и противоположных по знаку и находящихся на некотором расстоянии друг от друга.

Основные характеристики диполя:
Плечо диполя
![]() -
вектор по величине равный расстоянию
между зарядами и направленный от
отрицательного заряда к положительному.
-
вектор по величине равный расстоянию
между зарядами и направленный от
отрицательного заряда к положительному.
Электрический дипольный момент диполя
![]() .
.
Вектор
![]() приложен к центру диполя и направлен
от отрицательного заряда к положительному.
приложен к центру диполя и направлен
от отрицательного заряда к положительному.
![]() .
.
Вокруг диполя образуется электрическое поле, основными характеристиками которого являются напряженность (силовая характеристика) и потенциал и разность потенциалов (энергетические характеристики).
Напряженность электрического поля диполя.
По определению напряженность электрического
поля определяется как сила, действующая
со стороны поля на единичный пробный
заряд
![]() :
:
![]() .
.![]()
Пробным зарядом называется точечный положительный заряд, электрическое поле которого не искажает исследуемое электрическое поле.
По закону Кулона
![]() в вакууме и
в вакууме и![]() в среде.
в среде.
Если заряд
![]() - одиночный заряд, создающий поле, а
- одиночный заряд, создающий поле, а![]() ,
которым мы исследуем поле заряда
,
которым мы исследуем поле заряда![]() ,
то
,
то![]() ,
а
,
а![]() .
.
Графически электрическое поле изображается
с помощью силовых линий – линий,
касательные к которым совпадают с
вектором
![]() .
Для поля одиночного заряда электрическое
поле изображается так:
.
Для поля одиночного заряда электрическое
поле изображается так:
Если поле образуется несколькими зарядами, то напряженность в каждой точке общего поля определяется по принципу суперпозиции, который для поля образованного двумя зарядами диполя записывается как
![]() .
.
В скалярном виде напряженность находится так:
а) на оси диполя в точку, в которой
рассчитывается напряженность вносится
пробный заряд и определяется направления
векторов
![]() и
и![]() ,
которые направлены в одну сторону, затем
складываем их по правилу сложения
однонаправленных коллениарных векторов
,
которые направлены в одну сторону, затем
складываем их по правилу сложения
однонаправленных коллениарных векторов
![]()

б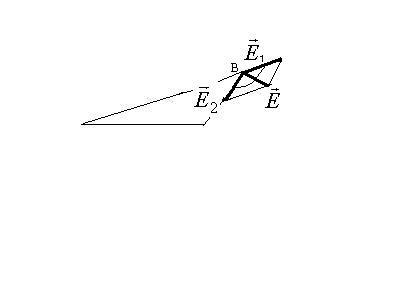 )
в произвольной точке В, на лежащей на
оси диполя:
)
в произвольной точке В, на лежащей на
оси диполя:
![]()
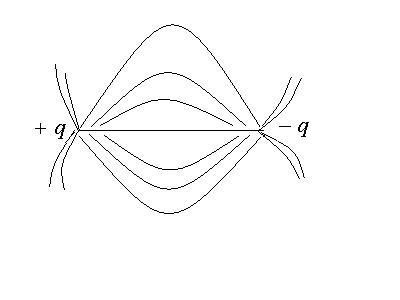
С помощью силовых линий электрическое поле диполя изображается
Потенциал. Разность потенциалов.
Напоминаю, что это энергетические характеристики электрического поля.
Потенциал электрического поля в любой его точке определяется как
![]() .
.![]()
и равен потенциальной энергии единичного заряда, внесенного в данную точку поля.
Если заряд
![]() переместить в поле из точки 1 в точку
2, то между этими точками возникает
разность потенциалов
переместить в поле из точки 1 в точку
2, то между этими точками возникает
разность потенциалов
![]() .
.
Смысл разности потенциалов: это работа электрического поля по перемещению заряда из одной точки в другую.
Потенциал поля также можно интерпретировать
через работую Если т.2 находится в
бесконечности, где поля нет (![]() ),
то
),
то![]() - это работа поля по перемещению заряда
из данной точки в бесконечность. Потенциал
поля, созданного одиночным зарядом
рассчитывается как
- это работа поля по перемещению заряда
из данной точки в бесконечность. Потенциал
поля, созданного одиночным зарядом
рассчитывается как![]() .
.
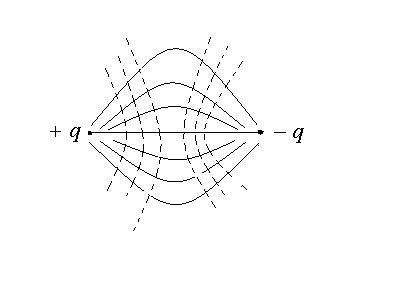
Поверхности, в каждой точке которой потенциалы поля одинаковы, называются эквипотенциальными поверхностями. В поле диполя потенциальные поверхности распределены следующим образом:
Потенциал поля, образованного несколькими
зарядами, рассчитывается по принципу
суперпозиции:
![]() .
.
а) Расчет потенциала в т. А, расположенной не на оси диполя:

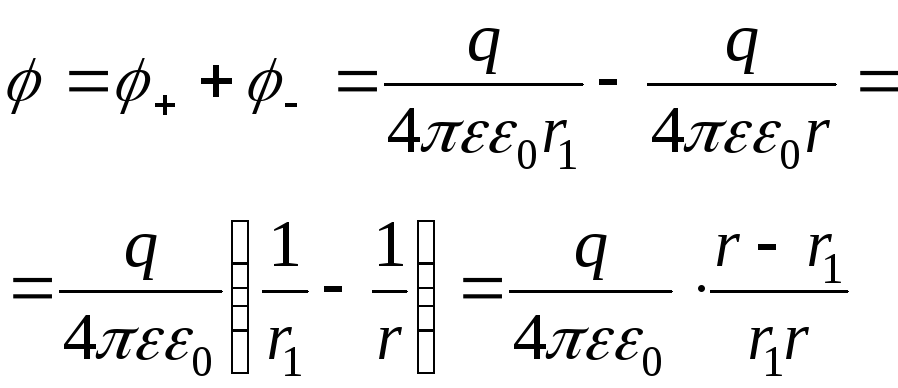
![]() найдем из треугольника (
найдем из треугольника (![]() ).
Очевидно,
).
Очевидно,![]() .
Поэтому
.
Поэтому![]() и
и![]() .
.
![]() .
.
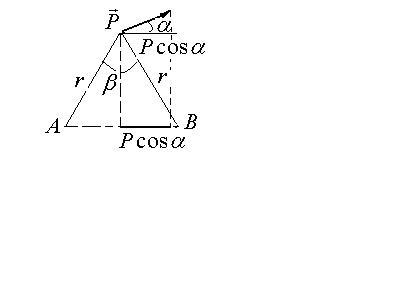
б) Между точками А и В, равноотстоящими
от диполя на расстоянии
![]()
(![]() )
разность потенциалов определяется как
(примем без доказательства, которое Вы
найдете в учебнике Ремизова)
)
разность потенциалов определяется как
(примем без доказательства, которое Вы
найдете в учебнике Ремизова)
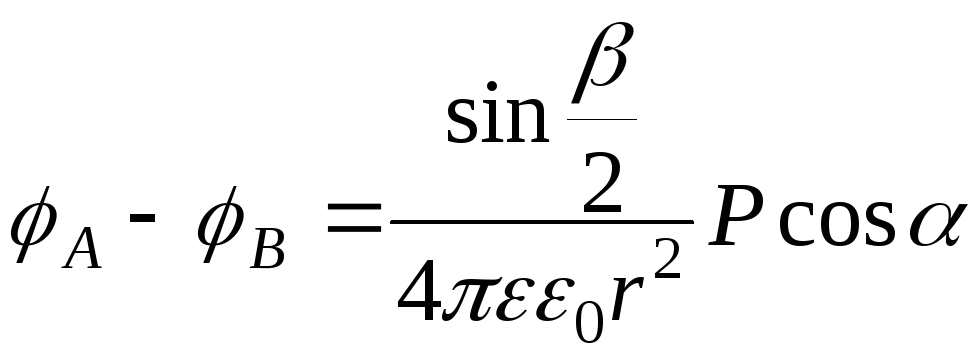 .
.
в) Можно показать, что если диполь
находится в центре равностороннего
треугольника, то разность потенциалов
между вершинами треугольника соотносятся
как проекции вектора
![]() на стороны этого треугольника (
на стороны этого треугольника (![]() ).
).
Диполь в электрическом поле.
а) В однородном поле напряженности
![]() .
.
![]()
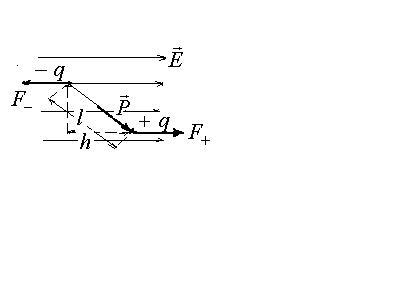
На каждый заряд диполя действует сила, соответственно
![]() .
.
Модули этих сил равны,
![]() ,
но направлены эти силы в противоположные
стороны, и, следовательно, создают
вращающий момент пары сил
,
но направлены эти силы в противоположные
стороны, и, следовательно, создают
вращающий момент пары сил
![]() ,
,
а это выражение есть векторное произведение
![]() .
.
Т.о. в однородном электрическом поле на
диполь действует момент сил, который
стремится установить диполь вдоль
силовых линий поля. Величина этого
момента зависит как от
![]() - характеристики диполя, так и от
- характеристики диполя, так и от![]() - характеристики поля, а также от
ориентации диполя.
- характеристики поля, а также от
ориентации диполя.
б) В неоднородном поле.
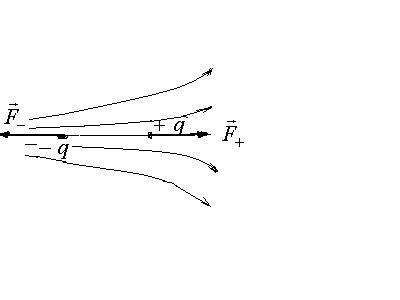
Если диполь расположен на силовой линии, то на его заряды действуют неравные силы:
![]() .
.
Здесь
![]() - напряженность поля в окрестности
положительного заряда,
- напряженность поля в окрестности
положительного заряда,![]() - напряженность поля в окрестности
отрицательного заряда. По густоте
силовых линий видим, что
- напряженность поля в окрестности
отрицательного заряда. По густоте
силовых линий видим, что![]() ,
т.к поде неоднородно.
,
т.к поде неоднородно.
Равнодействующая сил, действующих на диполь, есть
![]()
На протяженности диполя мерой
неоднородности поля служит величина
![]() - среднее изменение напряженности,
приходящееся на единицу длины диполя.
- среднее изменение напряженности,
приходящееся на единицу длины диполя.
![]() -
мера неоднородности поля вдоль
соответствующего направления поля.
Откуда
-
мера неоднородности поля вдоль
соответствующего направления поля.
Откуда![]() .
Следовательно,
.
Следовательно,
![]() ,
,
т.е.
![]() зависит от величины
зависит от величины![]() - характеристики диполя, так и от градиента
- характеристики диполя, так и от градиента![]() - характеристики поля. Под действием
силы
- характеристики поля. Под действием
силы![]() диполь втягивается в область большей
напряженности поля.
диполь втягивается в область большей
напряженности поля.
Если диполь находится не на силовой линии, то он и вращается, и втягивается в область больших значений напряженности поля.
Токовый диполь. Эквивалентный электрический генератор.
Рассмотренный нами диполь может сохраняться сколь угодно долго только в вакууме или идеальном диэлектрике. Однако нас диполь интересует в применении к объяснению электрических явлений в организме – электропроводящей среде. В такой среде диполь не сохраняется, т.к. свободные заряды среды, притягиваясь к противоположно заряженным полюсам диполя, либо экранируют его, либо нейтрализуют.
Т.к. электрическое поле, образующееся
в организме, не исчезает в процессе
жизнедеятельности, то необходима иная
модель диполя. Можно представить его
как электрический генератор с э.д.с.
![]() .
Если использовать его в цепи, то для
этой цепи закон Ома записывается как
.
Если использовать его в цепи, то для
этой цепи закон Ома записывается как![]() ,
,
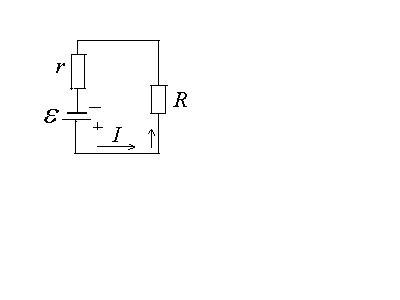
то сила тока зависит, от внешнего
сопротивления цепи
![]() .
Создавая модель диполя как источника
электрического поля, мы должны исключить
внешнее сопротивление. Примем
.
Создавая модель диполя как источника
электрического поля, мы должны исключить
внешнее сопротивление. Примем![]() ,
к клеммам источника диполь, или, иначе,
клеммы источника будем рассматривать
как полюса диполя, который в электропроводящей
среде будет сохраняться до тех пор, пока
функционирует источник тока.
,
к клеммам источника диполь, или, иначе,
клеммы источника будем рассматривать
как полюса диполя, который в электропроводящей
среде будет сохраняться до тех пор, пока
функционирует источник тока.
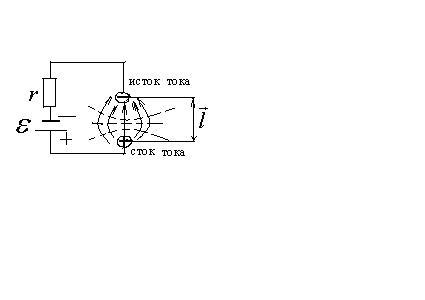
Отрицательный полюс назовем истоком тока, положительный – стоком тока.
Такую модель диполя называют токовым диполем.
Для токового диполя
![]() ,
следовательно, сила тока не зависит от
сопротивления среды, в которой он
находится
,
следовательно, сила тока не зависит от
сопротивления среды, в которой он
находится
![]() .
.
Дипольный момент токового диполя
![]() ,
,
где
![]() - плечо диполя, по модулю этот вектор
равен расстоянии. Между стоком и истоком
тока.
- плечо диполя, по модулю этот вектор
равен расстоянии. Между стоком и истоком
тока.
Потенциал поля токового диполя аналогичен
потенциалу электростатического диполя,
но электрические свойства среды в
формуле заменяются на её электропроводящие
свойства
![]()
![]() ,
,
![]() удельная электропроводность среды.
удельная электропроводность среды.
Суперпозиция токовых диполей называется эквивалентным электрическим генератором.
Электрокардиография
Итак, при функционировании органов и тканей, а также клеток в организме возникает электрическое поле, элементарным источником которого является диполь. Мы показали, что характеристики этого поля можно рассчитать, а это значит, что их можно измерить.
Метод регистрации разности потенциалов называют электрографией. В принципе он прост: достаточно двух электродов, накладываемых на пациента, гальванометра и усилителя. В зависимости от того, на каких органах регистрируется разность потенциалов электрографию классифицируют на ЭКГ – регистрация разности потенциалов при сокращении сердца; ЭЭГ – электроэнцефалография- регистрируется активность мозга; ЭМГ- регисрация активности мышц.
Мы рассмотрим электрокардиографию как наиболее распространенный метод.
За цикл работы сердца возбуждение
распространяется по различным отделам
его нервно-мышечного аппарата с
определенной последовательностью,
поэтому мгновенные значения результирующей
разности потенциалов за цикл работы
изменяется как по величине, так и по
расположению точек, между которыми они
имеют наибольшее значение. Из этих
значений наибольшей является разность
потенциалов между основанием и верхушкой
сердца в направ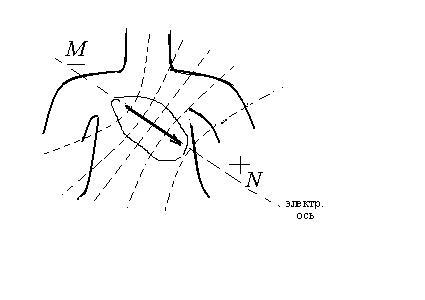 лении
электрической оси
лении
электрической оси![]() .
.
:

Кривая зависимости разности потенциалов от времени за время одного кардиоцикла называется электрокардиограммой
В основу электрокардиографии положена теория Эйнтховена:
1. сердце моделируется как источник разности потенциалов в виде токового диполя (эквивалентный электрический генератор);
2. диполь находится в однородной электропроводящей среде;
3. дипольный момент
![]() сердца образуется суперпозицией
дипольных моментов элементарных токовых
диполе, которые во множестве имеются в
возбужденном миакарде сердца
сердца образуется суперпозицией
дипольных моментов элементарных токовых
диполе, которые во множестве имеются в
возбужденном миакарде сердца
![]()
и называется интегральным дипольным вектором сердца (интеградьным дипольным моментом сердца)
Проекции эквипотенциальных поверхностей диполя на фронтальную поверхность тела показаны на рисунке штриховыми линиями;
дипольный момент сердца располагается во фронтальной плоскости тела;
5. точку приложения дипольного момента сердца можно считать постоянной – это нервный узел межпредсердной перегородки.
6. Связь между интегральным дипольным моментом и разностью
потенциалов определяется исходя из
наших прежних рассмотрений:
![]() .
.
При таком расположении вектора
![]() ,
как показано на нашем рисунке, разность
потенциалов наибольшая в направлении
электрической оси между верхушкой и
основанием сердца. На ЭКГ она соответствует
зубцу
,
как показано на нашем рисунке, разность
потенциалов наибольшая в направлении
электрической оси между верхушкой и
основанием сердца. На ЭКГ она соответствует
зубцу![]() .
.
Метод отведений Эйнтховена
Эйнтховен предложил при проведении
электрокардиографии измерять разность
потенциалов между каждыми двумя вершинами
равностороннего треугольника, построенного
симметрично относительно тела человека,
а центр теугольника совпадает с точкой
интегрального электрического вектора
сердца. Вершины этого треугольника
лежат на левом предплечье (ЛР), правом
предплечье (ПР) и левой ноге (ЛН). На
каждые две точки накладываются по
электроду, и между ними измеряется
разность потенциалов. Каждые две точки
наложения электродов называются
стандартными отведениями.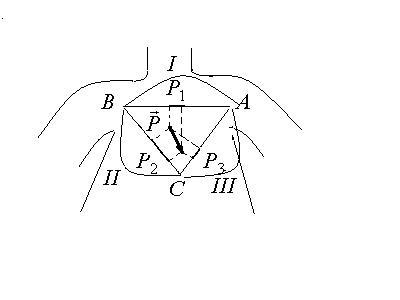
Отведения:
I– ЛР-ПР;
II– ЛН-ПР;
III- ЛН-ЛР.
Разности потенциалов между каждыми двумя точками пропорциональны проекциям дипольного момента на линию, соединяющую соответствующие точки:

Вектор-электрокардиография.
Точку приложения вектора
![]() можно считать постоянной (точка,
совпадающая с нервным узлом межпредсердной
перегородки), а конец вектора
можно считать постоянной (точка,
совпадающая с нервным узлом межпредсердной
перегородки), а конец вектора![]() за цикл работы описывает сложную
пространственную кривую. С помощью
осциллографа, используя его усилители,
можно наблюдать проекции этой
пространственной кривой на фронтальную,
горизонтальную и сагиттальную плоскости,
совмещенные с телом обследуемого.
за цикл работы описывает сложную
пространственную кривую. С помощью
осциллографа, используя его усилители,
можно наблюдать проекции этой
пространственной кривой на фронтальную,
горизонтальную и сагиттальную плоскости,
совмещенные с телом обследуемого.
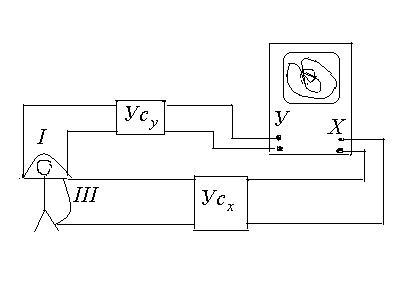
В плоскости при этом наблюдаются петли
![]() ,
образованные сложением взаимноперпендикулярных
колебаний мгновенных значений ЭКГ в
двух каких-либо отведениях (фигуры
Лиссажу)
,
образованные сложением взаимноперпендикулярных
колебаний мгновенных значений ЭКГ в
двух каких-либо отведениях (фигуры
Лиссажу)
Этот метод называется вектор-электрокардтографией,
а полученная кривая вектор-
электрокардиограммой (ВЭКГ).
Вектор-электрокардиограмма - кривая,
показывающая геометрическое место
точек, соответствующих положению конца
вектора
![]() за время одного цикла.
за время одного цикла.
Блок-схема осциллографа.
Электрокардиограмма регистрируется на приборе, называемом электрокардиографом. Он содержит следующие основные блоки:

Лекция 10
ЭЛЕКТРОДИНАМИКА
Электрические колебания.
Процессы, происходящие в идеальном
колебательном контуре.
Электромагнитные колебания–колебания величин заряда, силы тока, напряжения, эдс индукции.
Электромагнитные колебания создаются в закрытом колебательном контуре, который представляет собой электрическую цепь, содержащую катушку индуктивности и конденсатор.
М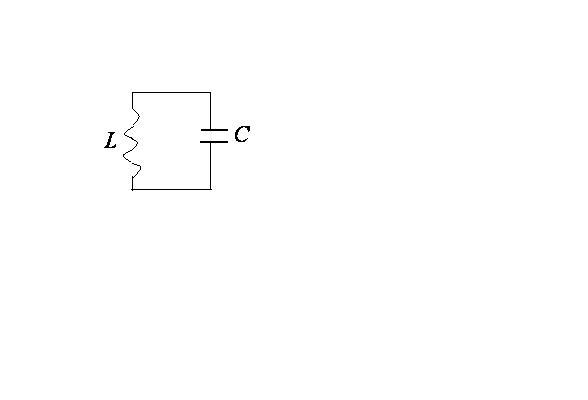 ы
начнем с изучения свободных (собственных)
колебаний, т.е. колебаний, которые
совершаются без внешнего воздействия
за счет первоначально накопленной
энергии.
ы
начнем с изучения свободных (собственных)
колебаний, т.е. колебаний, которые
совершаются без внешнего воздействия
за счет первоначально накопленной
энергии.
Рассмотрим идеальный колебательный
контур, т.е. контур, в котором активное
сопротивление
![]() (сопротивление проводов катушки) равно
нулю.
(сопротивление проводов катушки) равно
нулю.

Если переведем ключ в положение 1 , то
конденсатор зарядится от источника
тока так, что на его пластинах накопится
максимальный заряд
![]() (на одной пластине +, на другой-).Перебросим ключ в положение 2 и? будем
считать, что с этого момента времени
(на одной пластине +, на другой-).Перебросим ключ в положение 2 и? будем
считать, что с этого момента времени
![]() рассматриваем процессы, происходящие
в контуре.
рассматриваем процессы, происходящие
в контуре.