

метод_МПА
.pdf21
алкоголя корковая активность подавляется больше в правом полушарии.
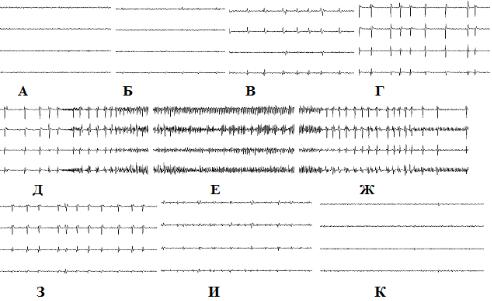
До введения раствора этанола ЭКоГ носила синхронизированно-десинхронизированный характер (рис 15). Через 2 минуты после введения раствора отмечалось увеличение представленности дельта активности в ЭКоГ. К 12 минуте после введения раствора в ЭКоГ увеличилась представленность медленноволновой активности. К 25 минуте после введения ВЭР отмечено снижение уровня медленноволновой активности в ЭКоГ и восстановление выраженности частых ритмов в ЭКоГ..
ФМПА показателей амплитуды ЭКоГ до и после применения ВЭР. До введения ВЭР (рис.17), в среднем за тридцатиминутный период наблюдения, показатели ФМПА амплитуд ритмов ЭКоГ выражалась статистически не значимыми величинами, а после введения ВЭР также в среднем за тридцатиминутный период наблюдения, показатели ФМПА амплитуд бета-2, бета-1 и альфа ритмов определялись статистически значимо измененными и положительными. Изменения показателей ФМПА амплитуды тета и дельта активности были статистически не значимы.
Рис. 15. Влияние внутрибрюшинного введения ВЭР на ЭКоГ крысы.
Обозначения: А – до введения этанола, Б - через 2 минуты после введения ВЭР, В - 12 минута после введения ВЭР, Г - 25 минута после введения ВЭР.

22
Показатели ФМПА по частоте ритмов ЭКоГ до введения ВЭР определялись статистически не значимыми, а после введения также определялись статистически значимо не измененными.
Показатели ФМПА индексов длительности бета-1, тета и дельта ритмов ЭКоГ до введения ВЭР определялись статистически не значимыми, а показатель индекса длительности бета-2 ритма был положительным и статистически значимым.
250 |
|
|
|
|
|
|
|
|
|
|
|
200 |
|
|
|
|
|
|
|
|
|
|
|
150 |
|
|
|
|
|
|
|
|
|
|
|
мкВ |
|
|
|
|
|
|
|
|
|
|
|
100 |
|
|
|
|
|
|
|
|
|
|
|
50 |
|
|
|
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
|
|
|
|
|
|
5 |
10 |
15 |
20 |
25 |
30 |
35 |
40 |
45 |
50 |
55 |
60 |
|
|
|
|
время (мин) |
|
|
|
|
|||
SB2A 
 SB1A
SB1A  SAA
SAA  STA
STA  SDA
SDA
Рис. 16. Динамика амплитуд ритмов ЭКоГ левого полушария до (5-30 мин) и после (35-60 мин) введения ВЭР.
После введения ВЭР показатель ФМПА индекса длительности бета-2 ритма статистически значимо инвертировал и выражался отрицательной величиной –6,75 2,20%. Отрицательной статистически значимой величиной выражался и показатель ФМПА индекса длительности дельта ритма - 8,65 1,61%. Показатели ФМПА индексов длительности альфа и тета ритмов были статистически значимо положительны
7,14 1,16% и 4,70 1,63% соотвтственно.
Моделируемые судорожные состояния являются достаточно интересной ситуацией для изучения изменений ФМПА.
Эпилептические приступы - основная мозговая дисфункция, затрагивающая 0.8 % людей. Многие из этих пациентов (20 %) являются фармакологически инкурабельны. Изучение патофизиологических механизмов эпилепсии, с целью прогнозирования приступов и их ингибиции, в этих

23
ситуациях, могло бы способствовать реабилитации этих больных.
15 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
% |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
α |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β2 β1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||
10 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
5 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
|
|
β1 |
α |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
-5 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
α |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
α |
|||||||||||
-10 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β1 |
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
-15 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
1 |
|
|
|
|
2 |
|
|
|
|
|
|
|
|
3 |
|
|
|
|
|
|
|
|
|
|
4 |
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||
Рис. 17. Диаграммы коэффициентов функциональной межполушарной асимметрии амплитуд ритмов 2-, 1-, -, -, и ЭКоГ, отводимой до (1, 2) и после (3, 4) применения ВЭР (1, 3) и сочетанного применения ВЭР и ПФв (2, 4)
Изучение механизмов эпилепсии требует использования биологических моделей, т.е. моделирования эпилептических признаков на животных. Для формирования биологических моделей генерализованных приступов эпилепсии применяют стимуляции таламуса, двусторонние корковые фокусы судорожной активности, интрацистернальные или системные введения пенициллина, гаммагидроксибутирата, внутрижелудочковое введение опиатов, генетические крысиные модели.
Биологическое моделирование эпилептической активности единовременно служит целям изучения структурно-функциональной организации головного мозга (Карлов А.А., 2002, Лобасюк Б.А., 1988) и изысканию корректных методов лечения эпилепсии.
Поэтому мы поставили перед собой задачу сформировать биологическую модель многоочагового эпилептического комплекс, на которой опыты можно было бы проводить в условиях свободного поведения животных.
24
Рис. 18. Эволюция судорожной активности сформированной внутрибрюшинным введением БП в дозе 300 000 МЕ и ПФ. А – до введения БП. Б – 280 сек после введения БП. В – 13 мин 50 сек сек после введения БП, 230 сек после введения ПФ. Г – 21 мин. 10 сек после введения БП. Д – 21 мин 33 сек после
Оказалось что при введении БП в дозе 300000 МЕ на 100 г массы тела животных в коре головного мозга крысы формируется модель многоочаговой корковой эпилепсии для которой характерны иктальные и интериктальные периоды (Спосіб моделювання осередкової кіркової епілепсії. Деклараційний патент на корисну модель 15.08.2005. Бюл. №8, Лобасюк Б.А., 2008).
После введения БП впервые судорожная активность возникала через 160-577 сек. от момента введения (рис. 18, Б) Из девяти крыс (4 контрольной и 5 опытной групп) у двух, первые судорожные разряды возникали в правом полушарии, а у шести в левом. У одной крысы возникновение судорожных потенциалов единовременно наблюдалось в обеих полушариях. Одновременно, с возникновением судорожной активности, в полушариях, не зависимо от стороны латерализации, ЭпА регистрировалась и в затылочном отведении. Позже всех ЭпА проявлялась в отведении фронтальной коры обоих полушарий, через 569 – 1188 сек.(9,48-19,8 минут) с момента введения БП (рис. 18, В). Формирование судорожной активности, в отведении
25
фронтальной коры во всех опытах, начиналось с веретенообразной активности. В этот период наблюдения во всех изучаемых регионах коры головного мозга регистрировались синхронно возникающие судорожные потенциалы – формировалась интериктальная судорожная активность (рис. 18, Г). Через 16-85 мин после введения БП у всех животных формировалась иктальная судорожная активность (16, 48, 57, 85 мин). Интервалы между иктальными разрядами составляли 2-50 мин. (рис. 18, Д, Е, Ж). В течение 3- 3,5 часов эволюции судорожной активности эпилептическая феноменология была относительно стабильна. В последующие периоды наблюдения амплитуды судорожных потенциалов уменьшались до их полного исчезновения (рис. 18, З, И, К).
Среднее время существования судорожной активности составило 313,25±14,94 минуты.
Динамика амплитуд (рис. 19) характеризовалась возрастанием величин судорожных потенциалов к 80 минуте наблюдения после введения БП от 579,11±46.13 мкВ в затылочной коре, до 1567,27±73,63 мкВ. в левом полушарии. Затем с 100-й минуты до 260 мин. наблюдался относительно стабильный период генерирования амплитуд судорожных потенциалов. К 340-й минуте наблюдения амплитуды судорожных потенциалов стремились к 0.
Изменения частоты генерации судорожной активности (рис. 20) носило волнообразный характер. Максимальные значения частот генерации судорожных потенциалов, регистрировались на 180 минуте после введения БП от 61,33±17,39 суд.пот/мин. во фронтальной коре, до 191,50±25,55 суд.пот/мин в затылочной коре.
Изменения показателей ФМПА ЭКоГ после исчезновения судорожной активности. До введения БП показатели ФМПА ( рис. 21) амплитуд бета-2, бета-1, альфа и тета ритмов ЭКоГ выражалась положительными значениями в пределах 4,00±0,93% – 9,81±2,89%, т.е. амплитуды этих ритмов были латерализованы в левое полушарие. Показатель ФМПА амплитуды дельта ритма выражался отрицательным значением, т.е. этот ритм по амплитуде был латерализоыван в правое полушарие. После окончания судорожной активности

26
показатели ФМПА амплитуды бета-2, бета-1, альфа и тета ритмов были отрицательны.
Рис. 19. Динамика амплитуд судорожных потенциалов вызванных внутрибрюшинным введением пенициллина в различных зонах коры головного мозга.
Обозначения: A1 - левое полушарие. A2 - правое полушарие. A3фронтальная кора, A4затылочная кора
Рис. 20. Динамика частоты генерации судорожных потенциалов вызванных внутрибрюшинным введением пенициллина в различных зонах коры головного мозга.

27
15 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
% |
α |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
10 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β1 |
|
|
θ |
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
5 |
|
|
β2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
δ |
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
-5 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
δ |
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
β2 |
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β1 |
α |
θ |
|||||
-10 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
-15 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
до введения пенициллина |
|
|
|
после исчезновения судорог |
||||||||||||||||
|
|
|
|
|
|
|
|||||||||||||||||
показатели
Рис. 21. Диаграммы коэффициентов функциональной межполушарной асимметрии амплитуд ритмов 2, 1, , θ и ЭКоГ до и после применения БП.
28
Литература
1.Адрианов О.С. Актуальные проблемы учения об организации функций мозга//Методологические аспекты науки о мозге.-М., 1983
2.Адрианов О.С. Осистемной организации церебральных функций. В кн.: Системный анализ механизмов поведения. М.:Наука, 1979. С.181-193.
3.Анохин П. К., Предисловие, в кн.: Г. Мэгун, Бодрствующий мозг, Изд-во иностр. лит., Москва (1961), с. 126
4.Ашмарин И. П., Кулашев А. П., Чепурнов С. А. Каскадные однонаправленные регуляторные процессы, осуществляемые короткоживущими пептидами (тиролиберин). Физиологический журнал СССР, 1989, т. 75, №5, 627-632
5.Бабенкова С.В. Клинические синдромы поражения правого полушария мозга при остром инсульте. М.: Медицина, 1971. 286. с.
6.Балабанов Ю.В., Варлинская Е. И. Химические факторы
формирования |
и |
|
фиксации |
устойчивых патологических состояний |
на |
моделях |
|
позиционной асимметрии // Вести. АМН СССР. 1981. - №
6. - С. 64-67.
7.Баллонов Л.Я., Деглин В.Л. Слух и речь доминантного и недоминантного полущария. Л.: Наука. 218 с.
8.Балонов Л.Я., Деглин В.Л. Слух и речь доминантного и недоминантного полушарий. Л.: Наука, 1976
9.Болдырева Г.Н. Электрическая активность мозга человека при поражении диэнцефальных и лимбических структур. М.: «Наука», 2000. 181 с.;
10.Болдырева Г.Н., Шарова Е. В., Добронравова И. С. «Роль регуляторных структур мозга в формировании ЭЭГ человека», Физиология человека, 26, № 5, 19-34 (2000)
11.Брагина Н.Н., Доброхотова Т.А. Функциональные асимметрии мозга человека. М: Медицина 1981, 288 с.
12.Вальдман А.В., Майский А.И., Кампов-Полевой А.Б., Тресков В.Г. Биологические основы поиска средств для
29
дифференцированной фармакотерапии алкоголизма//Вестню АМН СССР. –1988.-№3.-С.28-32.
13.Громов Л.О., Євтушенко О.А., Міпівкульова психофармакологія.-III Національний з ізд фармакологів України.Фармакологія крок у майбутнє. 17-20 жовтня 2006 р. Одеса. С.46.
14.Зиле Р.К., Одынец Т.Г., Клуша В.Е. Сравнительная оценка влияния пептидных гормонов, их фрагментов и соответствующтх аминокислот на содержание биогенных аминов мозга мышей- В кн: Люминесцентный анализ в медико-биологических исследованияхю Ринга. РМИ, 1980, с.90-95.
15.Клуша В.Е. Психотропные свойства гипофизарных пептидных гормонов. ― Хим.-фармац. журнал, 1978, № 3. с.3-5.
16.Клуша В.Е., Ауна З.П., Мышлякова Н.В., Макарова Н.А. Модели активных центров тканевых гормонов. ― Тез. докл. IV Всесоюз. съезда фармакологов „Фармакология ― здравоохранению”. Л., 1976. с.101.
17.Клуша В.Е., Муцениеце Р.К., Свиркис Ш.В. Фрагменты пептидных гормонов как возможные центральные регуляторные факторы.- В кн.: Нейрофармакология пептидов / Под ред. А.В. Вальдмана. М., НИИ фармакологии АМН СССР, 1982, с. 31-39.
18.Клуша В.Е., Свиркис Ш.В., Зиле Р.К. О центральных эффектах некоторых низкомолекулярных гормонов и их фрагментов. ― Изв. АН ЛатвССР, !978, № 8. с.100-113.
19.Коекина О.И., Гайдамакина А.М., Королева М.В., Возможности коррекции межплолушарной асимметрии ЭЭГ в процессе рефлексопрофилактики рецидивов алкоголизма //Журн.невропатол. и психиатр.- 1991, 2.- С.88-9
20.Лобасюк Б. А. Роль ретикулярной формации ствола мозга в механизмах коркового электрогенеза //Нейрофизиология/Neirofiziologiya, Т. 37, №; 1, с. 36 – 47.
21.Лобасюк Б.А. Анализ нейрофизиологических механизмов купирования многоочаговой корковой эпилепсии. Автореф. дис .канд. мед. наук, Новосибирск, 1988.- С.202
30
22.Ефремова Т. М., Труш В. Д. Динамика частотных характеристик биопотенциалов коры головного мозга кролика при становлении условного оборонительного рефлекса // Журнал высшей нервной деятельности им. И. П. Павлова.-1971.-т. 21.-в.5.- С. 963.
23.Ефремова Т. М., Труш В. Д. Электрофизиологическое исследование некоторых видов внутреннего торможения // Журнал высшей нервной деятельности им. И. П. Павлова,- 1974.- т. 24.- в.2.- С. 292.
24.Жадин М. Н., Бахарев Б. В. Анализ электрографических коррелятов привыкания и обучения с помощью методов распознавания образов,- В кн.: Физиологические и биохимические исследования памяти,- Пущино: НЦБИ АН
СССР, 1977.- С. 68.
25. Жадин М. Н., Бахарев Б. В., Муравьева Л. И. Электрофизиологические корреляты привыкания и выработки условного рефлекса //Журн. высш. нерв, деятельности им. И. П. Павлова.- 1977.- т. 27.- в.6,- С.
1173.
26.Лобасюк Б.А. Биологическое и математическое моделирование корковой эпилепсии внутрибрюшинным введением пенициллина// Вісник психіатрії та психофармакотерапії. 2008 № 1 (13) с.65-71.
27.Мосидзе В. М., Акбария К. К. О синдроме мозолистого тела // Журн. невропатол. и психиатр. 1973. № 3. С. 456461.
28.Мосидзе В. М., Мхеидэе О. А., Макашвили М. А. Асимметрия мозга человека. - Тбилиси: Мецниереба, - 1990. - 127 с.
29.Мельник Э.В. О природе болезней зависимости (алкоголизм, наркомания, «компьютеромания» и другие».- Одесса .-1988: изд. «Чорномор'я», 1998.-400 с.
30.Мосидзе В.М. Межполушарная интеграция //. Руководство по физиологии. Частная физиология нервной системы-Л., Наука, 1983, с. 690.
31.Рещикова Т.Н. Влияние алкоголя на латерализацию функций зрительного восприятия человека // ЖВНД. 1980. Т. 30. № 6. С. 1284–1287.