

Температурная рецепция
В коже, кроме болевых и механорецепторов, имеются и инкапсулированные терморецепторы, подразделяющиеся на две группы: тепловые (воспринимают тепло) и холодовые (воспринимают холод).
Холодовые рецепторы представлены колбами Краузе, тепловые — тельцами Руффини. В целом в коже насчитывают 250000 холодовых и 30000 тепловых рецепторов.
Колбы Краузе расположены в поверхностном слое дермы (на глубине 0,17 мм от поверхности кожи), имеют сферическую форму, окружены тонкостенной капсулой, нервные волокна перед вхождением внутрь колбы теряют миелиновую оболочку, ветвятся, переплетаются между собой (рис. 17. Б). Тельца Руффини располагаются в глубоких слоях дермы (0.3 мм от поверхности кожи). Преобладанием холодовых рецепторов и их поверхностным расположением можно объяснить парадоксальное первоначальное ощущение холода при погружении в теплую воду.
Терморецепторы располагаются в коже неравномерно, наибольшая плотность их отмечается на коже открытых частей тела (лицо, кисти рук), поэтому эти участки более чувствительны к температурным раздражителям по сравнению с участками тела, закрытыми одеждой. На некоторых участках кожи, воспринимающих тепло или холод, терморецепторы отсутствуют, поэтому полагают, что температурные раздражители могут восприниматься и свободными нервными окончаниями. Очень сильные тепловые или холодовые раздражители могут вызвать боль.
Температурные рецепторы имеются также в некоторых внутренних органах, гипоталамусе промежуточного мозга. При возбуждении температурных рецепторов нервные импульсы по афферентным путям поступают в центр терморегуляции гипоталамуса и рефлекторно включаются механизмы, обеспечивающие поддержание постоянной температуры тела.
При длительном воздействии тепла или холода происходит постепенная адаптация температурных рецепторов вследствие снижения их чувствительности к данным раздражителям.
Мышечная и суставная рецепция
В мышцах, сухожилиях, связках располагаются проприорецепторы (проприоцепторы), возбуждающиеся при растяжении и сокращении мышц. Импульсы от этих рецепторов, рецепторов кожи, глаз, вестибулярного аппарата передают в ЦНС информацию о степени сокращения или расслабления каждой мышцы, положении тела, его отдельных частей в пространстве, в результате осуществляется рефлекторная регуляция двигательных актов, контролируется правильность и точность движений.
По строению и функции проприоцепторы делятся на три группы:
-
Мышечные веретена;
-
Тельца Гольджи;
-
Тельца Пачини
Мышечные веретена имеют форму веретена, представляют собой рецепторы растяжения (механорецепторы). Они располагаются в мышце параллельно массе мышечных волокон и прикреплены чаще обоими концами к перимизию (оболочке пучка мышечных волокон) или иногда вторым концом к сухожилию мышцы. Каждое веретено содержит пучок тонких специализированных мышечных волокон, которые называются интрафузальными (лат. fusus — веретено), а мышечные волокна, составляющие основную массу мышцы — экстрафузальными волокнами..
Экстрафузальные волокна скелетной мышцы иннервируются α — эфферентными нервными волокнами (аксоны α — мотонейронов спинного мозга) и ответственны за развитие напряжения и сокращение мышцы.
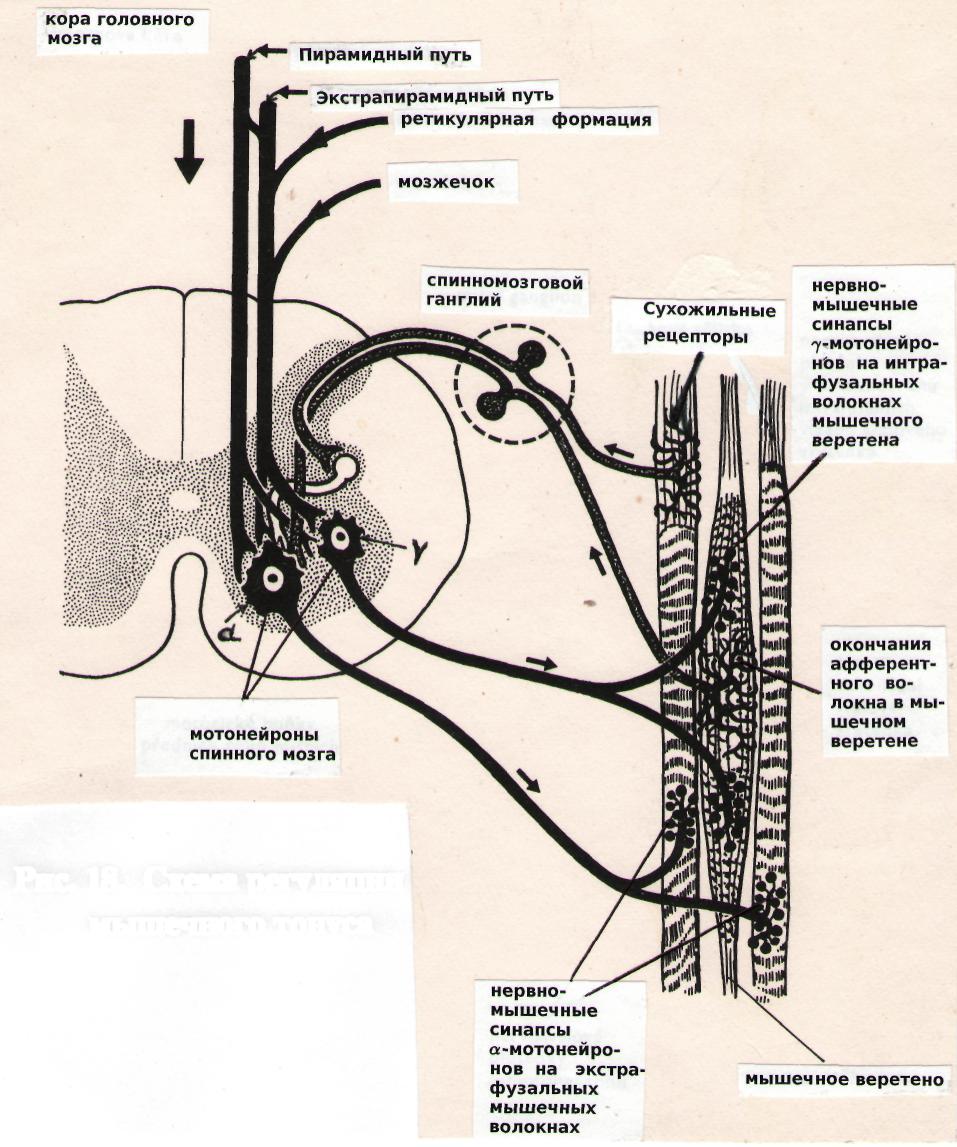
Рис. 18. Регуляция мышечного тонуса
Интрафузальные волокна состоят из центральной части (ядерной сумки) и двух периферических участков, способных к сокращению. Эти сокращающиеся участки интрафузальных волокон в отличие от экстрафузальных имеют менее выраженную поперечную исчерченность, иннервируются Υ — эфферентными нервными волокнами (аксоны Υ —мотонейронов спинного мозга). В центральной зоне мышечная ткань частично замещена соединительной и ретикулярной тканью, в ней располагаются спиралевидные нервные окончания толстых афферентных нервных волокон (рис. 18).
В естественных условиях существования основным раздражителем мышечных веретен является действующая на скелет сила земного притяжения, растягивающая скелетные мышцы, или сокращение мышцы-антагониста. Мышцы, ответственные за поддержание позы, должны в противовес силе тяжести удерживать скелет выпрямленным, Особенно значительному воздействию силы тяжести подвергаются мышцы-разгибатели. При растяжении мышц или их расслаблении происходит растяжение центральной части мышечного веретена, что вызывает возбуждение находящиеся в ней нервных окончаний. Потоки афферентных нервных импульсов от мышечных веретен поступают в ЦНС к α — мотонейронам спинного мозга, а от них по α — эфферентным нервным волокнам к экстрафузальным волокнам этой же мышцы, вызывая ее сокращение и торможение мышцы-антагониста (через тормозные интернейроны). При сокращении мышцы ослабевает растяжение мышечных веретен и импульсация от них в ЦНС прекращается.
Поток импульсов от мышечных веретен в ЦНС усиливается при сокращении периферических участков интрафузальных волокон веретена, так как происходит растяжение ядерной сумки и возбуждение рецепторов. Степень их сокращения регулируется импульсами, поступающим к ним по Υ — эфферентным нервным волокнам. Активность Υ — мотонейронов регулируется ретикулярной формацией среднего мозга.
Таким образом, мышечные веретена передают в ЦНС о состоянии мышц, их фактической длине и скорости ее изменения.
Сухожильные рецепторы Гольджи располагаются в зоне соединения мышечных волокон с сухожилием, образованы разветвлениями дендритов афферентных нейронов, которые свободно лежат или оплетают сухожильные и частично мышечные волокна. Эти окончания чувствительны к напряжению, развиваемому мышцей при ее сокращении. Импульсы, приходящие от сухожильных рецепторов, через тормозные интернейроны воздействуют на α — мотонейроны соответствующей мышцы, вызывая ее торможение. Интенсивность потока импульсов относительно пропорциональна силе сокращения мышцы, что позволяет рассматривать сухожильные рецепторы как источник информации о силе, развиваемой мышцей.
Тельца Пачини — инкапсулированные проприоцепторы (механорецепторы), расположенные в суставных сумках, фасциях, покрывающих мышцы, связках, Сокращение и расслабление мышц вызывает деформацию капсул, нервных окончаний, расположенных внутри их, и афферентный поток нервных импульсов поступает в ЦНС.
Количество мышечных веретен и сухожильных телец различно в различных мышцах. Чем более значимую функцию выполняют скелетные мышцы, тем больше они содержат мышечных веретен (например, мелкие мышцы кисти, особенно мышцы приводящей большой палец)
Информация от мышц и суставов поступает в спинной мозг, где она частично переключается на мотонейроны, а частично направляется по восходящим путям (пучки Голля и Бурдаха) в одноименные ядра продолговатого мозга. Здесь происходит переключение импульсов на нейроны второго порядка, образующие мощный бульбо-таламический пучок, получивший название медиальной петли, который заканчивается в ядрах таламуса. Отсюда берут начало пути, образованные нейронами третьего порядка, и оканчивающиеся в соматосенсорных зонах коры больших полушарий.