

Билеты_бф_экзамен_-_2019
.PDFстремящейся удержать вместе ионы с противоположными зарядами Равновесные потенциалы:
ENa=+35 … +65 мВ EК= -70 … -100 мВ
Состояние равновесия наступает в результате диффузии лишь очень небольшого количества ионов (по сравнению с их общим содержанием)
Зависимость мембранного потенциала (МП) нервного волокна каракатицы от наружной коннцентрации К (1) и величина МП, рассчитанная по уравнению Нернста для калиевого электрода (2)
Калиевый равновесный потенциал всегда больше (по абсолютному значению) реального потенциала покоя
Структуры мембраны, формирующие потенциал покоя:
потенциал управляемый Na-канал, Na/К – АТФаза, канал утечки, потенциал управляемый К-канал
Итоговая величина ПП, обусловленного переносом многих ионов может быть достаточно точно рассчитана по формуле Гольдмана
19.Электротон и локальный ответ, их сходство и различие
Электротон: Не способен распространяться по мембране Не изменяется ионная проницаемость мембраны Не способен к суммации
Амплитуда электротонического потенциала прямо пропорционально зависит от силы раздражителя Механизм пассивного электротонического потенциала:

Пассивный электротонический потенциал определяется только емкостными и резистивными свойствами клетки Электротонический потенциал не приводит к открытию потенциалуправляемых ионных
каналов. В этом случае емкостные и резистивные свойства мембраны клетки достаточно постоянны.
Емкость в основном определяется липидным бислоем Сопротивлением клетки зависит от сопротивления, которое определяется открытыми каналами утечки
Локальный ответ – при увеличении амплитуды подпороговых раздражителей от 0,5 до 0,9 пороговой величины развитие деполяризации мембраны происходит не прямолинейно, а по S-образной кривой.
Деполяризация продолжает нарастать и после прекращения раздражения, а затем сравнительно медленно исчезает Свойства:
-не способен распространяться по мембране
-изменяется ионная проницаемость
-способен к суммации
-амплитуда ЛО градуально зависит от силы раздражителя
20.Потенциал действия аксона: фазы и механизм формирования. Изменение ионной проницаемости мембраны во время генерации ПД
Потенциал действия разный для разных клеток различен: 0,5-1 мс – нервные клетки несколько мс – клетки скелетных мышц сотни мс – кардиомиоциты
Общая амплитуда – 100-120 мВ Овершут – около 30-50 мВ
ПП PK: PNa: PCl=1 : 0,04 : 0,45 ПД PK: PNa: PCl= 1 : 20 : 0,45
Чем больше ионов натрия, тем больше амплитуда ПД
Фаза деполяризации. Развитие ПД возможно только при действии раздражителей, которые вызывают деполяризацию клеточной мембраны. При деполяризации клеточной мембраны до критического уровня деполяризации (КУД) происходит лавинообразное открытие потенциалчувствительных Na+-каналов. Положительно заряженные ионы Na+ входят в клетку по градиенту концентрации (натриевый ток), в результате чего

мембранный потенциал очень быстро уменьшается до 0, а затем приобретает положительное значение. Явление изменения знака мембранного потенциала называют реверсией заряда мембраны.
Фаза быстрой и медленной реполяризации . В результате деполяризации мембраны происходит открытие потенциалчувствительных К+ -каналов. Положительно заряженные ионы К+ выходят из клетки по градиенту концентрации (калиевый ток), что приводит к восстановлению потенциала мембраны. В начале фазы интенсивность калиевого тока высока и реполяризация происходит быстро, к концу фазы интенсивность калиевого тока снижается и реполяризация замедляется.
Фаза гиперполяризации развивается за счет остаточного калиевого тока и за счет прямого электрогенного эффекта активировавшейся Na+ / K+ помпы.
Овершут – период времени, в течение которого мембранный потенциал имеет положительное значение.
Пороговый потенциал – разность между мембранным потенциалом покоя и критическим уровнем деполяризации. Величина порогового потенциала определяет возбудимость клетки – чем больше пороговый потенциал, тем меньше возбудимость клетки.
21.Na-канал: селективность, структура. Модель Хилле. Блокаторы, активаторы Na-канала. Роль натриевых каналов в генерации ПД
Модель Хилле
Основана на изучении проницаемости Na-канала для органических ионов
Na >ГИДРОКСИЛАМИН >ГИДРАЗИН >АММОНИЙ ФОРМАМИДИН ГУАНИДИН
МЕТИЛАММОНИЙ Радиус этих ионов примерно одинаков: 0,37-0,38 нм
Наибольшей способностью проникать через Na-канал обладают гидроксиламин и гидразин Метиламмоний несмотря на сходство размров с гидроксиламином и гидразином не проходит через канал
Причина: невозможность метильной группы образовывать водородные связи Вход в канал – прямоугольная пора 0,3*0,5 нм
Селективный фильтр в Na-канале содержит 8 атомов кислорода, по мнению других авторов селективный фильтр включает карбоксильную группу, которая создает сильное электрическое поле
Аминокислотные последовательности имеют четыре высокогомологичных домена, каждый из них содержит 6 гидрофобных участков
Блокаторы натриевого канала: ТЕТРОДОТОКСИН, САКСИТОКСИН, ДИНОФЛАГЕЛЛЯТЫ местные анестетики: прокаин
(новокаин), тетракаин, лидокаин, кокаин, морфин – блокируют натриевую проводимость посредством
взаимодействия с белками или мембранными липидами

Воздействия, устраняющие инактивацию натриевого канала: протеолитические ферменты, пептидные токсины (яд скорпиона, анемон), алколоидные нейротоксины (аконитин, батрахотоксин) - усиливают активацию
Вызывают активацию – вератридин, аконитин
Чем больше ионов натрия, тем больше амплитуда ПД
Фаза деполяризации. Развитие ПД возможно только при действии раздражителей, которые вызывают деполяризацию клеточной мембраны. При деполяризации клеточной мембраны до критического уровня деполяризации (КУД) происходит лавинообразное открытие потенциалчувствительных Na+-каналов. Положительно заряженные ионы Na+ входят в клетку по градиенту концентрации (натриевый ток), в результате чего мембранный потенциал очень быстро уменьшается до 0, а затем приобретает положительное значение. Явление изменения знака мембранного потенциала называют реверсией заряда мембраны.
22. К-канал: селективность, структура. Блокаторы К-канала. Роль выпрямляющих К-каналов
в генерации ПД
Проницаемость калиевого канала для одновалентных катионов не коррелирует с радиусом иона
PK : PRb : PCs : PLi : PNa= 1,0 : 0,91: 0,077: 0,018 : 0,01
Структура К-канала установлена с помощью тетраэтиламмония Калиевый канал асимметричен, имеет форму воронки. Диаметр выходного отверстия 0,3 нм, диаметр устья воронки 0,8 нм
Мак-Киннон и его коллеги расшифровали структуру и механизм функционирования нескольких бактериальных белков, каждый из которых формирует канал, проводящий ионы калия в ответ на изменение мембранного потенциала: работа калиевого канала, как лопастного
Блокаторы калиевого канала: антиаритмические препараты: амиодарон, хинидин, верапамил
Фаза быстрой и медленной реполяризации . В результате деполяризации мембраны происходит открытие потенциалчувствительных К+ -каналов. Положительно заряженные ионы К+ выходят из клетки по градиенту концентрации (калиевый ток), что приводит к восстановлению потенциала мембраны. В начале фазы интенсивность калиевого тока высока и реполяризация происходит быстро, к концу фазы интенсивность калиевого тока
снижается и реполяризация замедляется.
23.Na/К-АТФаза: строение и функционирование. Роль в генерации электрических потенциалов в электровозбудимых клетках.
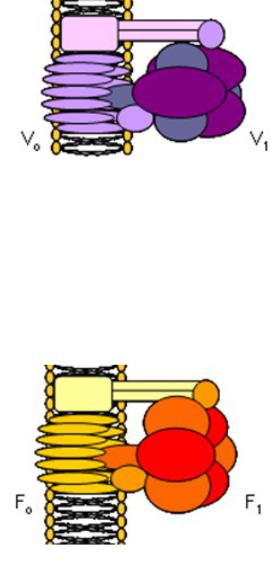
Существует 3 типа АТФ-аз: V-, F-, P-типа
V-типа: локализация: мембраны вакуолей дрожжей, тонопласты растений, лизосомы, эндосомы, секреторные гранулы
V0 (в мембране) – гидрофобная часть – участвует в транспорте протонов
V1 (в цитозоле) – водорастворимая часть, обладает каталитической активностью, катализирует как синтез, так и гидролиз АТФ Функции: переносят протоны, участвуют в транспорте анионов, аминокислот и репарации мембран при эндо- и экзоцитозе
Ингибиторы: нитраты, SH-реагенты, KSCN (тиоционат калия), ДЦКД (дициклогексилкарбодиимид)
F-типа: мембраны бактерий, хлоропластов и митохондрий F0 (в мембране) – гидрофобная часть, транслокация протонов
F1 – водорастворимая часть обладает каталитической активностью Ингибиторы: олигомицин, ДЦКД (дициклогексилкарбодиимид), ионы кадмия Функции: создание градиента протонов

P-типа: образование ФОСФОРИЛИРОВАННОГО продукта, который участвует в реакционном цикле
Примеры: Na/K-АТФаза, Са-АТФаза, Н-АТФаза плазматических мембран эукариотических клеток Ингибитор: ванадат-ион
Структура: состоит из 2 полипептидных цепей и Ингибитор: УАБАИН и др. сердечные гликозиды
Последовательность работы:
1.3Na связываются специфическим центром транслоказы
2.Изменение конформации транлоказы, вызванное присоединением 3Na, приводит к активации каталитической субъединицы и увеличению сродства активного центра к субстрату (АТФ). Протекает реакция аутофосфорилирования по карбоксильной группе аспарагиновой кислоты
3.Аутофосфорилирование изменяет заряд и конформацию транслоказы, она закрывается с внутренней стороны мембраны и открывается с наружной, уменьшается сродство к ионам натрия, и они диссоциируют от переносчика
4.Na/K-АТФаза, открытая с наружной стороны мембраны, имеет специфический центр связывания для 2К. Присоединение двух ионов калия к фосфорилированной транслоказе вызывает изменение конформации и появление аутофосфатазной активности. Протекает реакция аутодефосфорилирования
5.Дефосфорилирование изменяет заряд и конформацию транслоказы, она закрывается с наружной стороны мембраны и открывается с внутренней, уменьшается сродство к ионам калия и они диссоциируют от АТФазы
6.АТФ-аза возвращается в исходное состояние
Регуляция активности:
соотношение Na/К и содержание АТФ – факторы краткосрочной регуляции фосфорилирование протеинкиназами, что приводит к снижению активности – факторы

долгосрочной регуляции
24.Проведение возбуждения по нервному волокну. Кабельное уравнение. Особенности проведения импульса по миелинизированному нервному волокну.
Согласно закону Ома ток, текущий по осевому цилиндру:
dV/dx=-rii
I – ток, текущий по осевому цилиндру, х – расстояние от источника тока, ri – сопротивление аксоплазмы

Лямда – расстояние на котором величина Е уменьшается в V раз
Таким образом, входное сопротивление нервного волокна, от которого зависит скорость

распространения нервного импульса, также связана с диаметром этого волокна. Это означает, что с уменьшением диаметра нервного волокна входное сопротивление мембраны значительно возрастает, а при уменьшении – падает. Это явление определяет зависимость скорости проведения возбуждения от геометрических и функциональных неоднородностей нервных волокон. Резкое падение входного сопротивления мембраны будет подобно феномену ―перегрузка генератора в электрической цепи и замедлять проведение возбуждения вплоть до выпадения отдельных нервных импульсов и наоборот – ―облегчать проведение возбуждения при росте входного сопротивления в случае уменьшения диаметра нервного волокна.
Распространение потенциала действия вдоль мембраны нервного волокна будет неодинаково, и из-заэтого возникают локальные токи, посредством чего и происходит распространение возбуждения. В каждой точке генерируется новый ПД, однако, проводится не он, а возбуждение в виде локальных токов – пусковых механизмов для новых ПД.
Всостоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, т.к. липидная мембрана имеет высокое электрическое сопротивление. Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда. На границе возбужденного и невозбужденного участка начинает протекать электрический ток. Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения, в то время как ранее возбужденные участки возвращаются в состояние покоя. Таким образом, волна возбуждение охватывает все новые участки мембраны нервного волокна.
Вмиелинизированном нервном волокне участки мембраны, покрытые миелиновой оболочкой, являются невозбудимыми; возбуждение может возникать только участках мембраны, расположенных в области перехватов Ранвье. При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис 4.3 А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис 4.3 Б). Однако, в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис 4.3 В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.
25.Кабельное уравнение. Постоянная длинны. Постоянная времени.

26.Метод фиксации потенциала. Динамика ионных токов. Вольтамерная характеристики возбудимой мембраны.
Метод пэтч-кламп (метод локальной фиксации потенциала) введен в лабораторную практику Э. Неером и Б. Сакманом в 1976 г.
Клеточная мембрана формирует очень плотный контакт с поверхностью кончика микроэлектрода.
Между стеклом и мембранным фрагментом возникает контакт, имеющий гигаомное сопротивление.
В результате образуется электрически изолированный участок мембраны, и шум регистрирующего сигнала уменьшается на несколько порядков.
В силу конструкции системы ток I этого усилителя, проходя через сопротивление мембраны (Rм) изменяет МП так, что достигается равенство между МП и Е. При достаточном коэффициенте усиления усилителя и быстродействии системы МП практически фиксируется на уровне Е. При снижении Е и вслед за ним МП до КУД или более в мембране нервного волокна (кальмара) открываются потенциалозависимые натриевые и калиевые каналы, что порождает трансмембранные токи, которые и регистрируются на фоне поддерживаемого сниженного МП.