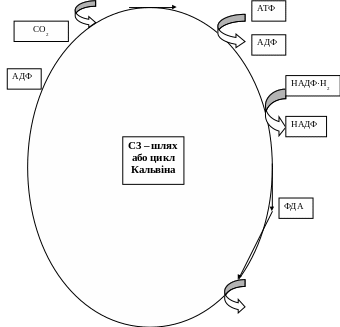
4. Цикл кальвіна
Послідовність
реакцій шляху С3
визначена Кальвіном і співробітниками
у 1946 – 1953 рр. У своїх дослідженнях Кальвін
використовував радіоактивний ізотоп
вуглецю
![]() і паперову хроматографію. Через 5 секунд
було виявлено перший продукт фотосинтезу,
ним виявилася фосфогліцеринова кислота
з трьома вуглецевими атомами. Звідси
друга назва циклу Кальвіна – шлях С3.
Кальвін з’ясував увесь ланцюг і
послідовність ферментативних перетворень
цих сполук і виділив при цьому три етапи:
карбоксилювання,
відновлення і регенерація,
які відбуваються у стромі хлоропластів.
Енергія, яка забезпечує цикл Кальвіна,
постачається у формі АТФ і НАДФ∙Н2,
утворених у світлових реакціях.
і паперову хроматографію. Через 5 секунд
було виявлено перший продукт фотосинтезу,
ним виявилася фосфогліцеринова кислота
з трьома вуглецевими атомами. Звідси
друга назва циклу Кальвіна – шлях С3.
Кальвін з’ясував увесь ланцюг і
послідовність ферментативних перетворень
цих сполук і виділив при цьому три етапи:
карбоксилювання,
відновлення і регенерація,
які відбуваються у стромі хлоропластів.
Енергія, яка забезпечує цикл Кальвіна,
постачається у формі АТФ і НАДФ∙Н2,
утворених у світлових реакціях.
Основна реакція циклу Кальвіна та, в якій бере участь СО2. вихідна сполука циклу Кальвіна – п’ятивуглецевий цукор з двома фосфатними групами – рибулозо-1,5-дифосфат.
Процес карбоксилювання починається тоді, коли СО2 входить у цикл і фіксується на рибулозо-1,5-дифосфаті. В результаті утворюється нестійкий проміжний продукт – цукор з шістьма атомами вуглецю, який відразу розкладається на дві молекули 3-фосфогліцеринової кислоти (ФГК). ФГК є першим продуктом фотосинтезу.
|
|
СН2ОР |
|
|
|
|
|
|
СН2ОР |
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
С |
|
О |
|
|
|
Н |
|
С |
|
ОН |
|
|
|
|
|
|
|
|
|
|
|
|
|
Н |
|
|
|
ОН |
|
|
|
НО |
|
С |
|
О |
|
|
|
|
|
С |
(С6) |
Н 2О |
|
|
+ |
|
|
Н |
|
С |
|
ОН |
|
|
|
НО |
|
С |
|
О |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
СН2ОР |
|
|
|
|
Н |
|
С |
|
ОН |
|
|
Рибулозо-1,5-дифосфат |
|
|
|
|
|
|
|
||||
|
|
|
|
|
СН2ОР |
|
||||||
|
|
|
|
|
|
3 фосфогліцеринова кислота |
||||||



 С
С
 О2
О2Цей найважливіший етап карбоксилювання каталізує фермент рибулозо-1,5-дифосфат-карбоксилаза-оксигеназа (РБФ-карбоксилаза). Вона знаходиться в стромі хлоропластів, її кількість становить 15% загальної кількості білка хлоропласту. Для здійснення каталітичної дії РБФ-карбоксилаза потребує присутності іонів Mg2+. Фермент складається з 8 великих і 8 малих субодиниць, які організовані в єдину структуру. Крім карбоксилазної активності фермент володіє також оксигеназною активністю, яку і проявляє у процесі фото дихання. Цей процес регулюється концентрацією кисню в повітрі.
Далі 3 - фосфогліцеринова кислота фосфорилюється з утворенням 1,3 – дифосфогліцеринової кислоти. Фосфорилювання відбувається за рахунок АТФ, синтезованого у світловій фазі фотосинтезу, за участю фермента фосфогліцераткінази.
|
|
СООН |
|
|
|
|
|
|
О |
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Н |
|
С |
|
ОН |
|
|
фосфогліцераткіназа |
|
|
С |
|
ОР |
|
|
|
|
|
|
+ |
АТФ |
|
|
|
|
|
|
|
|
|
СН2ОР |
|
|
|
|
Н |
|
С |
|
ОН |
||
|
3 - ФГК |
|
|
|
|
|
|
|
|||||
|
|
|
|
|
СН2ОР |
|
|||||||
|
|
|
|
|
|
|
1,3 – дифосфогліцеринова кислота |
||||||

+ АДФ
Наступний етап відновлення 1,3 – ДФГК за участю НАДФ∙Н2 – продукта світлової фази фотосинтезу, відновлюється до 3 – фосфогліцеринового альдегіду.
|
|
О |
|
|
|
|
|
|
СНО |
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
С |
|
ОР |
|
|
триозофосфат |
Н |
|
С |
|
ОН |
|
|
|
|
|
|
|
|
|
+ |
НАДФ∙Н2 |
|
|
|
|
|
|
+ |
НАДФ+ |
+ |
Н3РО4 |
Н |
|
С |
|
ОН |
|
|
дегідрогеназа |
|
|
СН2ОР |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
СН2ОР |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1,3 - ФГК |
|
|
3ФГА |
|
|
|
|
||||||||

Карбоксильна група СООН відновлюється до альдегідної групи СНО. Цю реакцію каталізує фермент гліцеральдегід–3–фосфат–дегідрогеназа. 3–фосфогліцеринів альдегід – це перший цукор, який утворюється при фотосинтезі. Як тільки утворився 3 – фосфогліцеринів альдегід, фотосинтез закінчується, бо усі подальші реакції відбуваються і в нефотосинтезуючих організмах. Утворені молекули 3-ФГА у циклі Кальвіна використовуються різними способами. 1/6 частина їх витрачається на синтез вуглеводів, серед яких переважають крохмаль і сахароза, 5/6 – йде на регенерацію рибулозодифосфату – первинного акцептора СО2. При використанні для синтезу вуглеводів молекула 3-ФГА ізомеризується в фосфодиоксиацетон. Цю реакцію каталізує фермент триозофосфатізомераза. Потім 2 триози-3-фосфогліцеринового альдегіду і фосфодиоксиацетону об’єднуються і утворюється цукор, яким є фруктозо-1,6-дифосфат, каталізує цю реакцію фермент фруктозодифосфатальдолаза. Утворений фруктозо-1,6-дифосфат дефосфорилюється і з’являється фруктозо-6-фосфат. Фосфатну групу відщеплює фермент фруктозодифосфат, а за допомогою ферменту глюзофосфатізомерази із фруктозо-6-фосфату утворюється глюкозофосфат. Таким чином з’являються моно фосфати глюкози і фруктози, з яких потім утворюється 2 основних вуглеводи – сахароза і крохмаль.
Для безперебійної фіксації СО2 необхідна постійна регенерація акцептора СО2 – рибулозодифосфату. Тому цикл Кальвіна замикає фаза регенерації. Регенерація – це складний цикл, в якому беруть участь цукрофосфати з 3,4,5,6,7 атомами вуглецю. У результаті багатьох реакцій утворюється рибулозо – 5 – фосфат. На його фосфорилювання витрачається решта АТФ, яке було синтезоване при фотофосфорилюванні у світловій фазі фотосинтезу. Регенерується вихідна сполука циклу Кальвіна рибулозо-1,5-дифосфат. По суті, рибулозо-1,5-дифосфат знову зазнає карбоксилювання і пройшовши ряд перетворень знову регенерується. У кожному оберті цього циклу 1 молекула СО2 відновлюється, а молекула рибулозодифосфату регенерується. Для утворення глюкози необхідно 6 обертів циклу з поглинанням 6 атомів вуглецю. Сумарне рівняння синтезу глюкози у циклі Кальвіна записується так:
6 СО2
+ 12НАДФ∙Н2
+ 18АТФ 1 глюкоза + 12НАДФ + 18АДФ +
6Н2О+18Фн
СО2
+ 12НАДФ∙Н2
+ 18АТФ 1 глюкоза + 12НАДФ + 18АДФ +
6Н2О+18Фн
Дослідження показали, що у циклі Кальвіна світло також необхідне, бо воно активізує ряд ферментів, серед них: рибулозо-дифосфаткарбоксилазу, гліцеральдегід-3-фосфатдегідрогеназу, фруктозодифосфатазу. Включення і виключення світла викликає переміщення протонів та іонів Мg2+ з внутрішнього простору тилакоїду у строму і навпаки. Ці зміни концентрації іонів у стромі допомагають активізувати вищезгадані ферменти. Цей шлях характерний для більшості рослин, у першу чергу дводольних.
Фруктозо-1,6-дифосфат
крохмаль
АТФ
Рибулозо-5-фосфат